2. Экологическая география генофонда и болезней населения Европейской России
|
Напомним, что главное, чего следует ожидать от исследования экологической геногеографии Восточной Европы - это ориентированного по географической широте распределения уровня заболеваемости населения, коль скоро именно такую ориентацию имеет 2-я, "экологическая", компонента изменчивости частот генов восточноевропейского генофонда. Поэтому, в дополнение к тому, что уже известно об этой компоненте из главы VI, представим ее в той трендовой форме, в которой она будет участвовать в проводимом здесь анализе (рис.ХIII-6, табл.ХIII-7).
2.1. География восточно-европейского генофонда. Результат корреляционного анализа тренда второй главной компоненты генофонда в географических координатах представлен в таблице XIII-7. Связь этого тренда с географической широтой оказывается по абсолютной величине выше, чем с долготой, что и позволяет говорить о преимущественно широтном направлении изменений генофонда по 2-й компоненте. Карта РС2.G читается двояко: в показываемой картой широтной зональности можно видеть отражение генетического влияния степных кочевых племен древности на население более северных территорий (этот аспект исторической геногеографии подробно рассмотрен в главе VI). Вместе с тем, в карте можно видеть отражение генетической адаптации восточноевропейского населения к природной зональности окружающей среды. Разночтения здесь нет, поскольку номады сами могли претерпеть генетическую, а не только хозяйственно-культурную, адаптацию к жизни в степях и распространять к северу не только сложившиеся в степном ландшафте навыки жизни, но и адаптивные комплексы своих генов. Тем не менее, разница между этими прочтениями существует, она принципиальна, но касается другого вопроса - о древности генетической адаптации и соответственно о древности PC2.G. 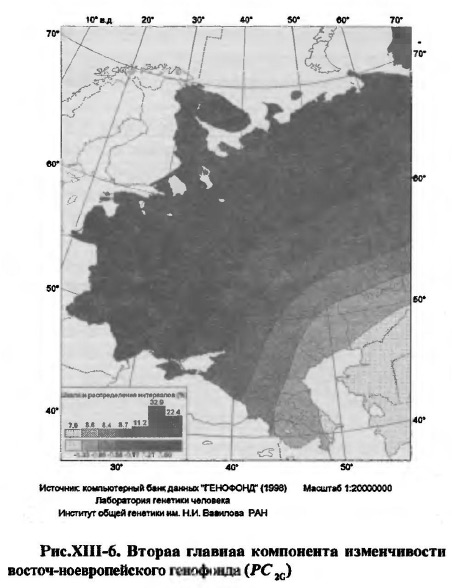

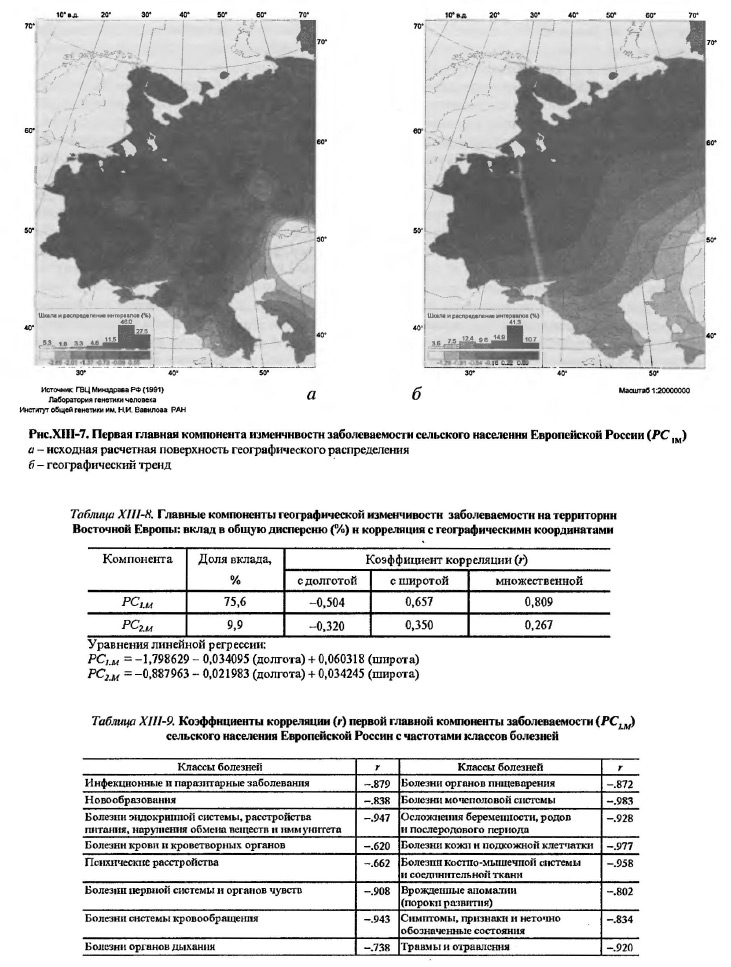
Генетическая адаптация населения к природной зональности окружающей среды могла длиться столько времени, сколько существует природная зональность. Выше уже были приведены археолого-географические и палеогеографические аргументы в пользу ее относительной, в геологическом масштабе времени, эволюционной молодости (поздний плейстоцен - голоцен), ниже мы коснемся этих аргументов вновь, применительно к особенностям ледникового периода в восточноевропейском регионе Северной Евразии. Что же касается проникновения адаптивных генных комплексов от степных кочевых народов в население лесостепной и лесной зон Восточной Европы, то эта возможность не могла появиться ранее, чем в степях возник номадизм (в энеолите-бронзе), а это делает возраст генетических адаптаций минимум на 5-10 тысячелетий моложе. Впрочем, номады эпох бронзы, раннего железа и средневековья могли дать дополнительный импульс генетической адаптации в том же - широтном - направлении географической дифференциации генофонда, которое определилось много ранее, в конце плейстоцена - начале голоцена, когда природа вступила в переживаемое и ныне очередное межледниковье. 2.2. География болезней сельского населения Европейской России. Исходя из нашей общей рабочей гипотезы о связи здоровья и болезней населения с его генофондом, следует ожидать, что широтная зональность обнаружится и в географии заболеваемости населения. Это ожидание можно подтвердить или опровергнуть картированием данных по заболеваемости сельского населения Европейской России. Сельскому населению отдано предпочтение перед городским по вполне очевидной причине более тесных его связей с природной средой, чем в городах, и большей стабильности этого населения, по сравнению с динамичными городскими популяциями, слабо или вовсе не воспроизводящими свой генофонд [Рычков, 1979]. Результаты анализа отражены в картах (рис.ХIII-7) и таблицах XIII-8 и XIII-9. На карте рисунка ХIII-7а представлена исходная расчетная поверхность географического распределения 1-й главной компоненты болезней 16 классов, а на карте рисунка ХIII-7б - географический тренд этой компоненты, выявленный при той же степени сглаживания в "плывущем окне", что и для генофонда (см. рис. XIII-6). Прогноз, основанный на рабочей гипотезе данного исследования, подтвердился: в географическом распределении болезней современного сельского населения Восточной Европы, действительно, обнаружена четко выраженная широтная зональность. При этом, изменение по географической широте оказалось свойственно не 2-й, как в генофонде, а 1-й главной компоненте (PC1.M), охватывающей 76 % общей дисперсии частот заболеваний 16 нозологических групп. Столь высокий удельный вес PC1.M означает высокую корреляцию частот всех или большинства нозологических групп с географической широтой местности, или, что практически то же самое, - с собственной первой главной компонентой (см. табл.ХIII-9). Действительно, связь всех нозологических групп заболеваний с PC1.M высока (-0.983< r < -0.620), а для половины из них особенно высока (r < -0.9): болезни нервной системы и органов чувств (r = -0.908), травмы и отравления (r = -0.920), осложнения беременности, родов и послеродового периода (r = -0.928), болезни системы кровообращения (r = -0.943), болезни эндокринной системы, расстройства питания, нарушения обмена веществ и иммунитета (r = -0.947), болезни костно-мышечной системы и соединительной ткани (r = -0.958), болезни кожи и подкожной клетчатки (r = -0.977), болезни мочеполовой системы (r = -0.983) (см. табл.ХIII-9). Хотя полностью исключить нельзя, но все же маловероятно, что столь высокая корреляция вызвана прямым воздействием фактора географической широты и связанных с ним природно-климатических особенностей местности на состояние здоровья современного сельского населения. Это маловероятно хотя бы потому, что население защищено от такого прямого воздействия экраном культуры народа, в том числе культуры хозяйственной жизни в сельских условиях. Как отмечалось выше, хозяйственно-культурные типы населения явно конгруэнтны к природной зональности. Таким образом, в случае PC1.M и РС2.G с мы имеем две несопоставимых по вкладу в изменчивость (76% и 12%, соответственно) главных компоненты двух, на первый взгляд независимых, явлений - заболеваемости и генофонда, которые, тем не менее, имеют однотипные географические распределения, характеризующиеся высокой (r = 0.945) положительной корреляцией (табл.ХIII-10). 
Как видно из данных таблицы XIII-9, практически всем 16-ти нозологическим группам заболеваний свойственны высокие и очень высокие отрицательные корреляции с PC1.M, указывающие на то, что географическое распределение PC1.M формируется действием некоего мощного фактора, перекрывающего частные отклонения в географической изменчивости частот болезней отдельных независимых нозологических групп. Иными словами, при всей разности этиологии и патогенеза анализируемых групп болезней, географическая изменчивость уровня заболеваемости по каждой из этих групп определяется общим для всех них фактором. Природа этого фактора может быть связана с одним из двух явлений; имеющих однотипную географию: широтная зональность природной среды или же широтная зональность в географической изменчивости самого генофонда. Русское национальное сознание настолько уверенно отождествляет природу средней полосы России как оптимальную для жизни, что трудно представить плавно перетекающую из одного широтного состояния в другое зональность русской природы как источник каких-то особых угроз и адаптивных требований к генофонду. Легче представить обратное: наш генофонд так давно и тщательно вписан в природную зональность России, что она стала восприниматься национальным сознанием как важнейшая часть жизненного и духовного благосостояния народа и его генофонда. Иными словами, дело не только в том, что трудно допустить одинаковое и прямое влияние природной зональности на заболевания разных нозологических групп, но прежде всего в том, что путем прошедших генетических адаптации природная зональность оказалась включенной в широтную зональность генофонда и из фактора внешней среды превратилась в фактор внутренней, генотипической, среды организма человека. В географической корреляции распределений генофонда и заболеваний незримо присутствует история формирования этих распределений. Для генофонда постановка вопроса об истории формирования главных компонент его изменчивости вполне закономерна, поскольку в этой изменчивости материализовано историческое время [Рычков, 1984,1986,1989, 1991а]. Гораздо менее обосновано, на первый взгляд, предполагать, что и за географией болезней стоит их история. Тем не менее, такое предположение вполне соответствует представлениям теоретической медицины о закономерностях развития болезней, которые кроются не только в глубинах онтогенеза, но и в глубинах эволюционной истории человека [Давыдовский, 1962]. Применим это положение теоретической медицины не к общему ходу эволюции человека, а к конкретному ходу истории в исследуемом восточноевропейском регионе Северной Евразии. Ориентируясь по карте РС1.M (см. рис.ХIII-7б) и по данным о корреляциях (см. табл.ХIII-9), следует предполагать единое направление географического хода этой истории для болезней совершенно различных этиологии и патогенеза. Однотипность связи распределения разных нозологических групп болезней с географической широтой с полной определенностью указывает именно на единый фактор истории, обусловивший современную географию болезней сельского населения Европейской России. Подчеркнем еще один важный момент в географии заболеваемости: высокая положительная корреляция PC1.M с географической широтой и высокие отрицательные корреляции болезней всех нозологических групп с собственной PC1.M означают, что низкий уровень заболеваемости свойственен сельскому населению северных и северозападных областей Восточной Европы, а высокий - южным и юго-восточным. Этот вывод из корреляционного анализа, на первый взгляд, парадоксален, поскольку противоречит обычным представлениям об ухудшении природных условий жизни в более высоких широтах и о более благоприятных для жизни южных областях. Более того, этот результат картографо-статистического анализа оказывается в явном противоречии с предложенной демографами региональной типологией здоровья населения России [Население России, 1993, с.54]. Согласно демографической типологии, уровень здоровья сельского населения европейской части страны, оцениваемый индексом потенциальной жизнеспособности, оказывается низким и очень низким в северных областях и в областях средней полосы и увеличивается в южных. Причина противоположности демографического и генетического заключений о географии здоровья, думается, связана с разными источниками анализируемых данных. Наш источник - данные о болезнях, редкость которых (большое дополнение частоты заболевания до 1) рассматривается как показатель здоровья по схеме "не болен - здоров". Источник, на основе которого демографами оценивается медико-санитарное благополучие и тип здоровья - это данные о младенческой смертности и средней ожидаемой продолжительности жизни. Для генетика на основании совокупности известных ему самых разноплановых данных о "развертывании" свойств генотипа в онтогенезе естественно предполагать, что в географическом распределении смертности, особенно младенческой, могут существовать особые закономерности, которые не сводятся к геногеографии болезней взрослого населения. Для демографа такое предположение может выглядеть избыточным. Каким же все-таки образом могло возникнуть показываемое нашими картами географическое распределение уровня заболеваемости, обратное "здравому смыслу"? Ответ на этот вопрос будет дан ниже, когда мы обратимся к истокам формирования широтной зональности в географии болезней, а здесь рассмотрим еще одну возможную причину отмеченного несоответствия - возрастной состав населения. Напомним, что картируемые данные о болезнях относятся к возрастному интервалу 18 и более лет; в этом интервале всегда численно преобладает группа до 59 лет (до 54 лет у женщин). Если доля этой возрастной группы, допустим, меняется по широте, то наблюдаемое на карте уменьшение уровня заболеваемости к северу может оказаться следствием более молодого возрастного состава населения более высоких широт за счет меньшей доли лиц, доживающих до пожилого возраста1. Карта, обозначаемая далее AG>59 (рис.XIII-8a), подтверждает факт убывания к северу доли возрастной группы >59 лет. Правда, эта карта показывает иной тип распределения, чем карты PC1.M заболеваемости и РС2.G с генофонда. Тем не менее, корреляция между картами PC1.M и AG>59 (табл.ХIII-11) весьма существенна (r = 0.551, η2 = 0.344) и указывает на 34-процентное влияние возрастного состава населения на уровень его заболеваемости; кроме того, карта возрастного состава в такой же мере коррелируется с картой генофонда (r = 0.578, η2 = 0.389), что ставит дополнительную проблему возрастного отбора, происходящего в генофонде. 
Однако, проведенный анализ частных корреляций между картами показывает (табл.XIII-12), что основная интересующая нас связь географической изменчивости заболеваемости (2) с генофондом (1) практически не зависит от возрастного состава (3) населения (коэффициент корреляции r1,2-3 = 0.920 остается высоким даже с исключением влияния возраста). Таким образом, связь заболеваемости с возрастом населения возникает через генофонд, состав которого меняется естественным отбором в разных возрастных группах населения. Из обусловленных возрастом генетических свойств населения обратим внимание на уровень гетерозиготности населения. Об изменении уровня гетерозиготности по возрастным группам сообщалось неоднократно и, в частности, подробно исследовано в коренном населении Чукотки [Соловенчук, 1996]. Однако, общее освещение проблемы появилось лишь в последнее время, показав для широчайшего спектра видов растительных и животных организмов универсальную роль уровня гетерозиготности организмов в определении сроков полового созревания и продолжительности жизни [Алтухов, 1996]. Исходя из этого исследования, можем предположить, что отрицательная связь уровня гетерозиготности с продолжительностью жизни косвенно может проявить себя в связи возрастного состава населения с уровнем гетерозиготности. 
Теоретически, гетерозиготность как квадратичная функция генных частот тем меньше будет повторять общую географию генофонда, чем более полиаллельны локусы, включенные в исследование генофонда. В нашем исследовании среднее число аллелей на локус составляет 2.94 ~ 3, то есть, "средний локус" трехаллелен, и география его гетерозиготности может отличаться от географии генофонда. Средняя на локус, ожидаемая гетерозиготность Ht = 0.3559 в Восточной Европе убывает с юго-запада (max = 0.3681) на северо-восток (min = 0.3319) (рис.ХIII-8 б), то есть, точно также, как и доля старшей возрастной группы. Географическая корреляция между ними довольно велика (r = 0.661), причем, влияние географии гетерозиготности на географию возрастного состава (67%) выше, чем обратное (60%). Географическое изменение уровня гетерозиготности, действительно, как это можно было предположить [Алтухов, 1996], влияет на изменение возрастного состава популяции и не связано с изменениями ни генофонда (r = -0.129), ни заболеваемости населения (r = -0.067). Проведя анализ частных корреляций, получим зависимость "гетерозиготность - возраст популяции", избавленную от опосредствующих влияний генофонда и болезней (см. табл.ХIII-12). Как видим, частная корреляция карт среднего уровня гетерозиготности (4) и возрастного состава населения (3) особенно велика (r3,4-1,2 = 0.918) на фоне исключенных влияний генофонда (1) и уровня заболеваемости (2). Дальнейшее развитие этой темы выходит за рамки данной главы. Как видно из таблицы XIII-12, корреляция карт возрастного состава (3) и гетерозиготности (4) населения мало сказывается на основной интересующей нас связи между географическими изменениями генофонда (1) и заболеваемости (2). От парной корреляции r1,2 = 0.945 через частные корреляции первого порядка r1,2-3 = 0.920 и r1,2-4 = 0.947 связь заболеваемости с генофондом лишь несколько уменьшается при исключении влияния возраста и гетерозиготности (r1,2-3,4 = 0.864), оставаясь достаточно высокой. Таким образом, остается непреложным фактом, что компоненты PC1.M заболеваемости и РС2С генофонда, независимые по источникам и природе стоящих за ними данных, имеют однотипные сильно скоррелированные географические распределения (см. табл.ХIII-12). Как уже отмечалось выше, в географической корреляции генофонда и заболеваний незримо присутствует история их формирования. Необходимо обратиться к истокам формирования широтной зональности в географии болезней. Эти истоки, учитывая высокую географическую корреляцию между PC1.M болезней и РС2.G с генофонда (см. табл.XIII-10), следует искать там же, где и истоки генофонда - в конце плейстоцена - начале голоцена [Развитие ландшафтов..., 1993]. Будем исходить из того, что особенности материальной культуры отражали ход культурной адаптации палеолитических охотников и собирателей к природным условиям и что эта культурная адаптация в Восточной Европе выражена гораздо более отчетливо, чем на остальных пространствах Северной Евразии, не испытавших покровного оледенения. 1 Авторы признательны проф. Г.Д.Засухииой за указание на это обстоятельство. |
загрузка...