Глава VI. Ю.Г. Рычков, Е.В. Балановская, С.Д. Нурбаев, Ю.В. Шнейдер. Историческая геногеография Восточной Европы
Постановка задачиГеногеография народонаселения способна напрямую связывать генетическую действительность современных поколений с древностью, какой бы глубокой она ни была. Ограничение, существующее для такого нового подхода к изучению прошлого, состоит в том, что геногеография, основываясь на изученных к настоящему времени генах, может в прошлом обнаруживать лишь предшественников этих именно генов. Но в этом ограничении заключено и определенное преимущество нового подхода: в прошлом времени для геногеографии открывается не вообще все, что в нем существовало, а лишь то, что генетически "проросло" в современность, то есть, те группы древнего населения, которым исторические обстоятельства благоприятствовали в передаче их генов в генофонд современных поколений. Вот почему вполне обосновано ставить проблему исторической геногеографии и разрабатывать новый источник исторической информации. В этой связи важно отметить, что основатель геногеографии А.С.Серебровский [1928, 1930] настаивал на том, что она является исторической, а не биологической наукой. Это утверждение можно понять лишь так, что геногеография использует биологические метки - гены, но исследует с их помощью географическое отражение исторического процесса. Исторический смысл геногеографии видел и Н.И.Вавилов, не только писавший об этом [Вавилов, 1968], не только избравший геногеографический подход к исследованию происхождения культурных растений, но и следивший за археологическими открытиями неолитических земледельческих культур, в том числе и трипольской культуры, раскопки которой он посещал, и приглашавший археологов для сообщений в Институт генетики. Обычно геногеография понимается как география генов, изучаемая с помощью их географического картирования. Каждый полиморфный ген, изменчивый в частоте встречаемости, может быть картирован, и каждой такой карте может быть предложено историческое прочтение, поскольку географическая изменчивость частоты гена есть результат ее изменений во времени. Но, все же, для исторической геногеографии мы считаем этот путь малоплодотворным из-за необозримости такого природного явления как полиморфизм генов. Этим создается почва для неограниченного числа версий исторического процесса на одной и той же картируемой территории, поскольку множество независимых полиморфных генов отразят историю населения этой территории во множестве независимых типов своего географического распределения. Более того, разнообразие этих распределений, а с ним и разнообразие исторических трактовок, будет множиться с открытием все новых и новых генов. География каждого гена в отдельности может передать лишь некоторую часть исторической информации. По нашим оценкам [Рычков, Ящук, 1980], в геногеографической картине отдельного гена содержится в среднем около трети исторических шумов, не связанных с этнической историей населения, что создает достаточно большие искажения в исторической интерпретации карты. Тем не менее, постановка вопроса об исторической геногеографии вполне правомочна. Но для этого нужно отойти от элементарного представления о геногеографии как географии генных частот и смириться с тем, что карта любого отдельно взятого гена отразит историю населения в неминуемо искаженном виде. Как избежать этих искажений? Для этого необходимо подняться на более высокую ступень геногеографического анализа, где возможно использовать замечательное свойство генов, вовлеченных в исторический процесс. Если в эволюционном процессе гены "ведут себя" по-разному, поскольку эволюция и есть перестройка генного состава популяции в соответствии с меняющимися условиями среды, то для исторического процесса все гены одинаковы, можно сказать, одинаково безразличны, не имеют отношения ни к содержанию, ни к направлению этого процесса. Гены вовлекаются в него не сами по себе, а через своих носителей, вовлекаются как щепки в поток и позволяют следить за потоком истории сколь угодно долгое время, будучи сами вечными в масштабе исторического времени. С ходом истории может как угодно измениться частота гена, но не самый ген. Каждый отдельно взятый ген (если оставим в стороне его нуклеотидную последовательность) сегодня такой же, каким был во все прошедшие времена с самой зари человеческой истории; это все равно, как если бы археолог мог располагать одним и тем же руководящим признаком для всех культур, всех историко-культурных областей и всех археологических эпох, начиная с раннего палеолита. Надо только найти способ следить сразу за множеством генов, коль скоро перед историей они все равны. Заменим их отслеживание во времени наблюдением в пространстве, то есть геногеографией, но такой, которая позволяла бы создавать карту не отдельных, а многих генов одновременно - синтетическую карту. Чем больше генов в нее включено, тем более надежно такая карта передаст географию исторического процесса на картируемой территории. Такова суть реализованного в этой работе подхода к исторической геногеографии. Что же касается метода, то ниже он будет указан лишь в самом общем виде, который позволяют рамки работы, но прежде остановимся на истории изучения генофонда населения Восточной Европы. История изучения восточноевропейского генофондаИстория исследования генофонда населения Восточной Европы насчитывает более ста лет. Еще в последней четверти XIX века русский морской военврач А.В Любинский [1885] изучал дальтонизм - генетически обусловленное нарушение цветового зрения - у новобранцев-матросов из различных губерний России. Начало изучения в России полиморфизма групп крови АВО было положено одновременно Н.К.Кольцовым [1922], проводившим исследования этих групп крови у населения Москвы и Подмосковья, и В.В.Бунаком [1924], изучившим распределение групп крови у белорусов Могилевщины. Большой вклад в накопление данных о генофонде России был внесен в результате сотрудничества медиков и антропологов в работе Комиссии по изучению кровяных групп (1927-1931 гг.) под руководством проф. В.Я.Рубашкина. Интенсивной и систематической была деятельность Б.В.Вишневского, руководившего Ленинградским бюро комиссии. Усилиями антропологов и врачей группы крови изучались у населения Поволжья (Б.В.Вишневский, Г.И.Петров, И.Н.Елистратов, Л.М.Геселевич), севера (Н.Н.Чебоксаров) и центра России (Г.Ф.Дебец), Кавказа (ЕЛ.Семенская). Кроме столичных научных центров, работа по изучению групп крови проводилась и в Приуралье, и в Сибири - в Перми (Б.В.Парин), в Томске (А.Б.Вагнер). В 30-е годы в программы исследования генофонда были включены изучение полиморфизма вкусовосприятия фенилтиомочевины (РТС) и групп крови MN, в конце 40-х годов - групп крови Rhesus, а затем и целый ряд новых систем групп крови. После трагического для отечественной генетики 1948 года собственно популяционно-генетические исследования населения СССР были приостановлены, однако, накопление генетической информации продолжалось усилиями медиков [Бронникова, 1947; Дробышева, 1948; Умнова, 1951]. Полевые исследования с определением групп крови для характеристики населения были возобновлены антропологами в 1957-1959 гг. В 1959 г. на кафедре антропологии МГУ была учреждена постоянная Сибирская генетико-антропологическая экспедиция. На базе названной экспедиции был выполнен и ряд популяционно-генетических исследований народов Европейской части СССР: белорусов [Микулич, 1972], саамов [Хазанова, 1973], литовцев [Гаусик, 1975]. В 60-е годы этапное исследование по геногеографии Восточной Европы было проведено В.В.Бунаком, выделившим ряд крупных геногеографических зон на основе данных станций переливания крови о группах крови АВО. Популяционной единицей исследования в этой работе выступала административная область, и геногеографические зоны охватывали группы областей [Бунак, 1969]. В 60-е же годы зародилось и выросло в самостоятельное направление исследование генетико-биохимического полиморфизма в популяциях человека [Спицын, 1967, 1968, 1985]. В эти годы большой вклад в накопление информации о генофонде населения продолжали вносить медики Института судебной медицины (А.К.Аржелас, М.А.Бронникова, М.И.Потапов), Московской городской станции переливания крови (А.Г.Башлай), Центрального института гематологии и переливания крови (М.А.Умнова), Ленинградского института переливания крови (Д.А.Серова) и многих других медицинских учреждений. В конце 60-х - начале 70-х годов в России появились первые специализированные генетические учреждения и лаборатории, разрабатывающие различные аспекты генетики человека. В 1967 г. был создан Институт медицинской генетики АМН СССР, в работах которого существенное место заняли популяционные исследования населения, в том числе севера Европейской части России [Пасеков, Ревазов, 1975; Ревазов и др., 1975, 1979]. Исследования генофонда населения СССР, проводившиеся ранее на кафедре антропологии МГУ, были продолжены и развиты с созданием в Институте общей генетики АН СССР лаборатории генетики человека, укомплектованной антропологами и руководимой проф. Ю.Г.Рычковым. В рамках работ этой лаборатории были охарактеризованы генофонды всех народов Кавказа (И.С.Насидзе, З.П.Инасаридзе, Л.А.Шен- гелия), а также большинства народов Европейской части России (Ю.В.Шнейдер). Материал и методыМатериал Материалом для данного исследования послужила информация о частотах генов в популяциях коренного населения, накопленная в созданном в лаборатории генетики человека ИОГен РАН Банке данных ГЕНОФОНД. Блок информации о генофонде населения Восточной Европы содержит данные о частотах 100 аллелей 36 генных локусов из 170 опубликованных с 1885 по 1995 гг. литературных источников, а также из архивов лаборатории генетики человека ИОГен РАН. Представление о степени изученности отдельных локусов в населении Восточной Европы (количество изученных по каждому локусу этносов и. популяций, общая чиссленность обследованных, вариабельность генных частот) можно получить из таблицы VI-1. Географическая локализация этих данных в виде карты опорных точек, на которых основываются представленные в работе результаты картирования генофонда, показана на рисунке VI-1. Генетические характеристики каждого из народов Восточной Европы по наиболее хорошо изученному генному локусу - системе групп крови AB0 - представлены в таблице VI-2. Как видно из таблиц, накопленной к настоящему времени информации о генофонде населения Восточной Европы вполне достаточно для его исследования с использованием методов геногеографии. Методы создания и анализа карт Принципиальная методическая новизна данной работы состоит в том, что в ней применена компьютерная технология создания электронных геногеографических карт, разработанная в лаборатории генетики человека Института общей генетики. Подробно методы построения геногеографических карт изложены в работах Е.В.Балановской и соавторов [1994а; 19946; 1995]. Напомним лишь, что в основе любой компьютерной карты лежит ее цифровая модель (ЦМ) - числовая двумерная матрица значений признака. Координатами этой матрицы служат узлы равномерной сетки, густо покрывающей карту. Значения матрицы рассчитываются с помощью интер- и экстра- поляционной процедуры по исходным значениям картируемого признака в изученных популяциях. Исходные данные картируемых генетических признаков распределены географически нерегулярно в любом исследуемом регионе. Для вычисления регулярно расположенных значений цифровой модели карты в узлах равномерной сетки применен метод средневзвешенной интерполяции по нерегулярным исходным точкам. В данной работе были применены следующие параметры интерполяционной процедуры: - показатель степени локального сглаживающего полинома 1; - радиус влияния весовой функции (расстояния от исходной точки до интерполируемого узла равномерной сетки) 10 см., в масштабе приводимых в работе карт этот радиус охватывает популяции в пределах 2000 км; - показатель степени весовой функции 6; Получив таким образом матрицы (ЦМ) картируемых признаков- карты распространения в Восточной Европе частот каждого из 100 картируемых генов - мы можем использовать их для любого вида одно- и многомерного статистического анализа. В таком анализе каждая карта выступает как числовая матрица и потому допускает любые преобразования [Балановская и др., 19946; Балановская, Нурбаев, 1995; Koshel, Musin, 1994]. В результате преобразования получается новая матрица, визуализируя которую мы получаем принципиально новую карту - не исходных признаков, а результата их картографо-статистического преобразования. 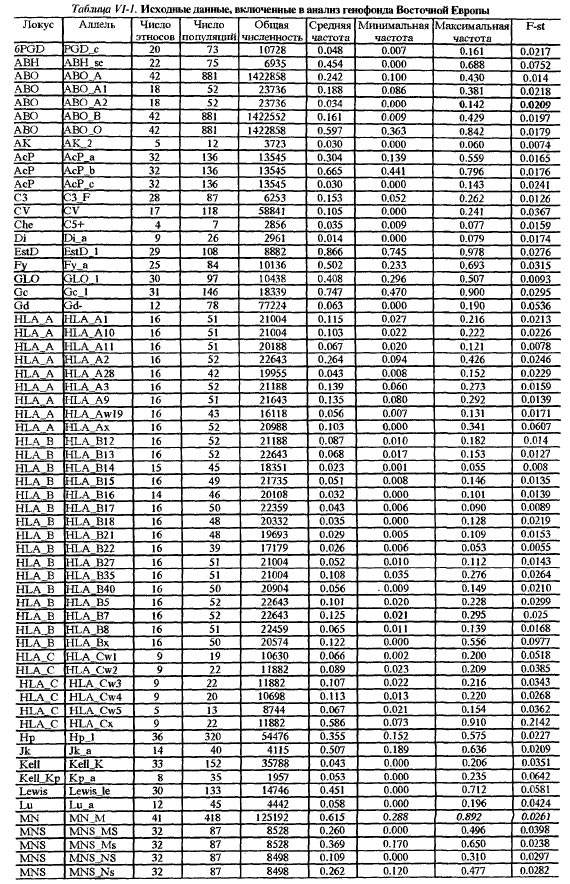
Таблица VI-1. Исходные данные, включенные в анализ генофонда Восточной Европы 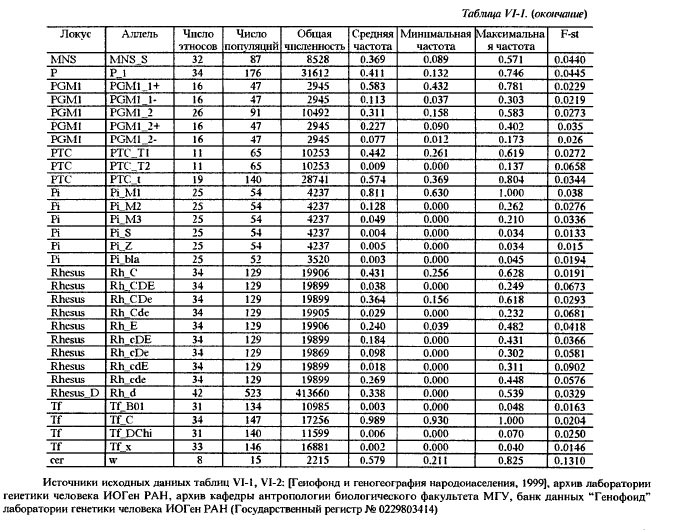
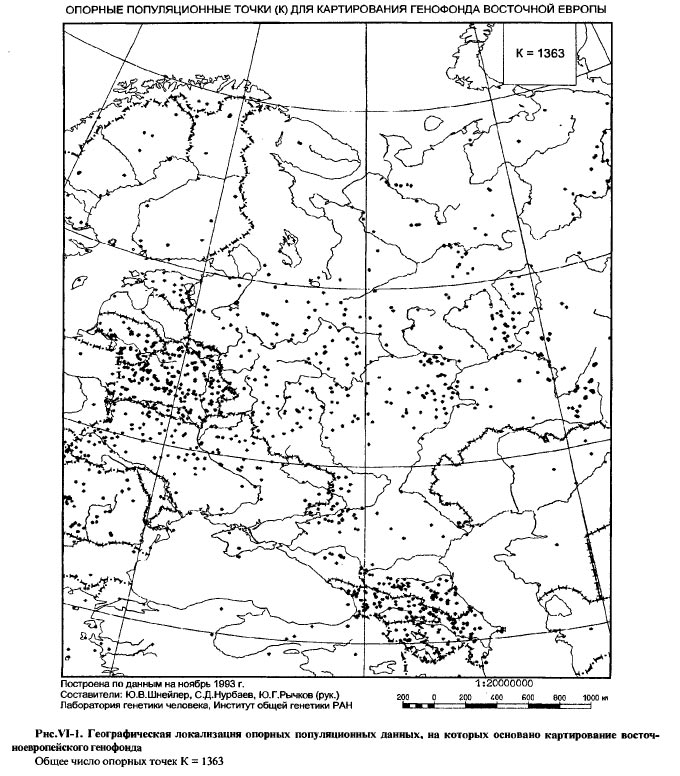
ОПОРНЫЕ ПОПУЛЯЦИОННЫЕ ТОЧКИ (К) ДЛЯ КАРТИРОВАНИЯ ГЕНОФОНДА ВОСТОЧНОЙ ЕВРОПЫ 
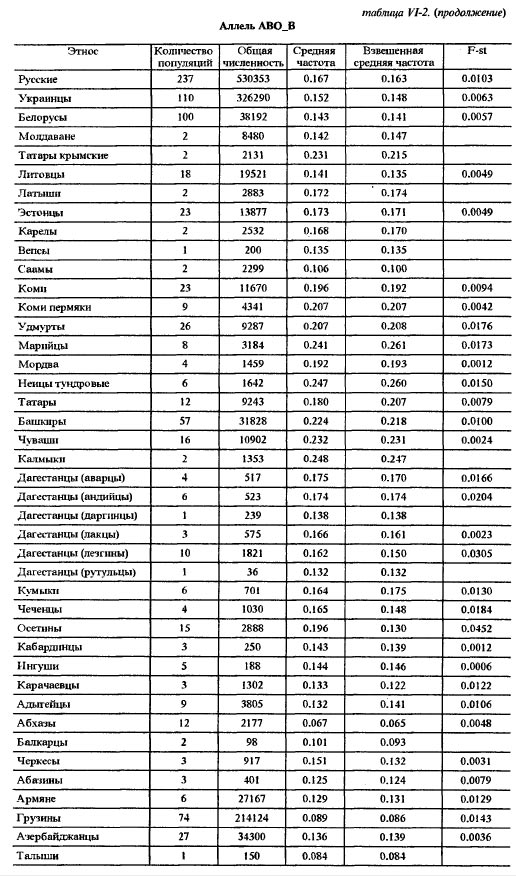
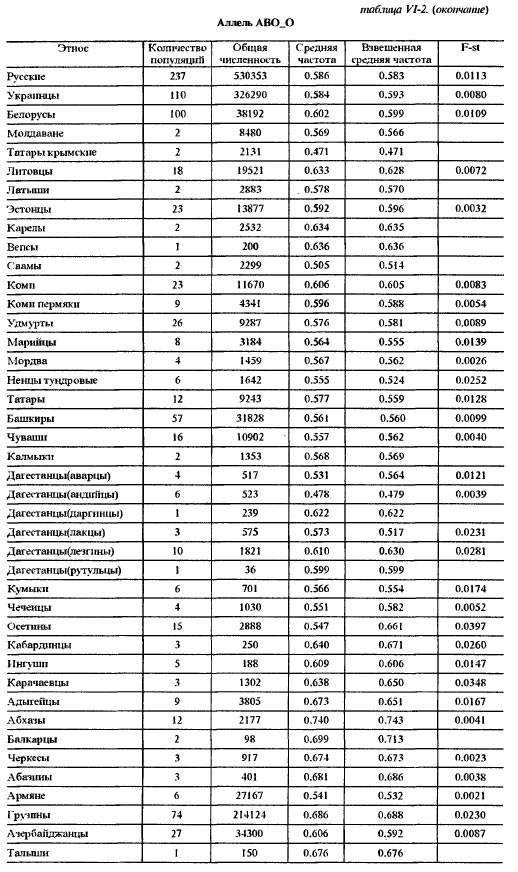
Таблица VI-2. Исходные данные по генам системы групп крови АВО Аллель АВО_А В данной работе использованы два картографо-статистических метода - 1) расчета и картирования главных компонент; 2) расчета и картирования генетических расстояний. Первый метод получил широкое распространение [Menozzi et al., 1978; Piazza et al., 1981; Рычков, Балановская, 1992] и не нуждается в дополнительных пояснениях. Для картирования генетических расстояний был применен специально разработанный метод, учитывающий особенности картографической информации и ориентированный на нее. Метод впервые использован в антропогеографическом исследовании следов летописных славян в современном русском населении [Рычков, Балановская, 1988] и в компьютерной форме реализован в исследовании по геногеографии монголов [Рычков и др., 1990; Балановская и др., 1990]. Существо метода состоит в том, что картируются не собственные значения тех или иных антропологических, генетических и других признаков, а величины их отклонений от некоторых заданных значений. Такими заданными значениями могут быть средние величины признаков какой-либо этнической группы или в пределах какой-либо территории, причем, картироваться могут отклонения от этих средних далеко за пределами данной этнической группы или территории. Привлекательность данного метода в том, что используемая "средняя" частота позволяет обнаружить в терминах генетических расстояний "близость" или "отдаленность" картируемых районов от заданных среднерегиональных значений. При этом выявляются территории распространения равновесных (близких к заданному среднему значению) частот генов для данного региона, а также области экстремумов. В данной работе в качестве заданных "средних" частот использованы средние частоты всех картированных генов в квадрате карты, вмещающем исторический ареал русского народа. Естественно, что этот квадрат захватывает малые ареалы исторических соседей и предшественников русских - балтов и восточных финнов. Но именно в силу малости их ареалов это слабо сказывается на оценке свойственных русским средних частот генов, так как частота каждого гена рассчитывается как взвешенная средняя, и весом выступает площадь территории. Выявленные территории указывают не только на субстратные слои генофонда и на направления миграций генов. Они, как и карты главных компонент, помогают увидеть обобщенный образ генофонда, прочитать историю его развития и взаимодействия - как с другими генофондами, так и с природной средой. Приводимые в этой работе карты трех первых главных компонент охватывают 42% совокупной изменчивости 100 карт отдельных генов. Эти три карты должны, по сути примененного метода, показать три взаимонезависимых (ортогональных друг другу) типа географического распределения множества генов. Интерпретация главных компонент, в общем случае, непростая задача. Мы предлагаем именно историческую интерпретацию, потому что лишь исторический процесс может воздействовать на множество независимых генов одинаковым образом и вызвать их согласованное перемещение по территории. Методически трудный момент здесь в том, что степень этой согласованности, похоже, может быть одинаковой при кратком, но мощном импульсе, испытанном генофондом, и при долговременном, но слабом воздействии. Поскольку гены способны сохранить и передать воздействия обоих типов, постольку синтетическая карта может представлять синтез не только разных географий разных генов, но и синтез разных времен, в которые имело место одно и то же географическое направление исторического процесса. Вместе с тем, будем иметь в виду, что устоявшиеся представления о силе исторических импульсов в тот или иной период времени, могут расходиться с результатом геногеографического анализа. Он имеет дело с генетическими последствиями импульсов, которые лишь приблизительно или вовсе не могут быть оценены по имеющимся историческим источникам. Например, история позволяет оценить степень воздействия монголо-татарского ига на развитие русского и других народов Восточной Европы, но вопрос о его воздействии на генофонд остается открытым, и не исключено, что геногеография внесет коррективы в представления о его масштабах. В заключение методического раздела несколько слов о том, как читать геногеографическую карту главной компоненты. Значения компоненты на карте ранжированы по величине. Трактуя компоненту как результат воздействия исторического процесса на генофонд, не следует принимать малую величину компоненты за малое историческое воздействие. И минимум, и максимум значений компоненты на карте есть два полюса, где исторические импульсы наиболее интенсивны, но географически противоположно направлены: ведь компоненты выделены именно по географическому направлению. Так, циклон и антициклон, соответственно, с низким и высоким атмосферным давлением, - оба сказываются на погоде, а их устойчивые области - на климате. По этой аналогии синтетические карты главных компонент могут быть прочитаны как карты исторического климата, и даже рассчитана для него "Роза ветров истории" [Рычков, Балановская, 1992]. Еще одна важная деталь, помогающая чтению карты - это форма изолинии. Если в какой-то части карты изолиния имеет выпуклую форму, то направление выпуклости указывает на географическое направление давления на генофонд, производившего в нем изменения именно в этом направлении. При вогнутой форме изолинии область ее вогнутости есть результат действия встречной силы изменения генофонда. Поэтому между выпуклым и вогнутым ходом изолиний всегда находится, даже если (из-за свойств шкалы) она не изображена на карте, прямая изолиния, указывающая на область равновесия между противоположно направленными силами, формирующими ту или иную из главных компонент генофонда. Обратимся же к синтетическим картам населения Восточной Европы, чтобы увидеть географию исторического процесса, запечатленную в геногеографии современного населения. Результаты и обсуждениеЗапад - Восток, 1-ая главная компонента изменчивости восточноевропейского генофонда На первый взгляд, карта 1-ой компоненты (рис. VI-2) отражает европеоидно-монголоидное деление территории, с границей, проходящей от Каспия через Поволжье до Белого моря, и с явным территориальным преобладанием (см. гистограмму площадей) высоких - восточных - значений этой компоненты над низкими - западными. В целом эта картина достаточно соответствует устоявшимся представлениям об антропологическом составе населения Восточной Европы [Алексеев, 1969; Алексеева, 1973]. Но структура генетического рельефа с выраженно дугообразным ходом изолиний в западно-центральной и южной частях карты и с почти меридиональным ходом изолиний в восточной части карты указывают на более сложное устройство генофонда, несводимое к антропологически классическому соотношению европеоидности и монголоидности в населении. Клин, образуемый серией изолиний в направлении с юго-запада на северо-восток, может быть понят как геногеографическое отображение истории проникновения европейских генов в генофонд населения Восточной Европы и постепенного замещения ими генов автохтонного и азиатского происхождения. Таким образом эта карта говорит не просто о соотношении в населении европеоидности и монголоидности в антропологическом смысле, но именно о проникновении в пределы Восточной Европы европейского генофонда. По отношению к нему, генофонды населения Кавказа и Малой Азии, с одной стороны, Прибалтики, Карелии, Фенно-Скандии - с другой, не говоря уже о Поволжье, Приуралье, Казахстане и Средней Азии - все они, безотносительно к уровню своей монголоидности, на этой карте оказываются одинаково чужеродными, принадлежащими к другому - не европейскому - миру, к иным историко-географичесим странам. Таким образом, карта 1-ой компоненты изображает генетические импульсы, идущие из Европы вместе с теми историко-культурными воздействиями, которые сознанием самих восточно-европейцев воспринимаются как влияния "Запада". 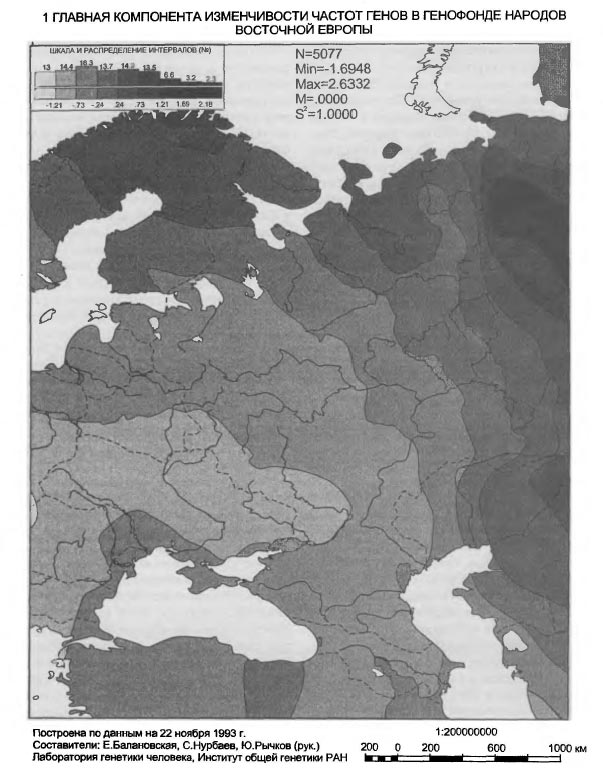
1 ГЛАВНАЯ КОМПОНЕНТА ИЗМЕНЧИВОСТИ ЧАСТОТ ГЕНОВ В ГЕНОФОНДЕ НАРОДОВ ВОСТОЧНОЙ ЕВРОПЫ К какому времени эти западные воздействия можно отнести? В карте потенциально отображено любое время, в котором европейский импульс был основным в развитии населения Восточной Европы. Например, черты такой именно географии заселения Восточной Европы впервые обозначились еще в позднем палеолите в Вилендорф-Павлово-Костенковском палеолитическом комплексе [Софер, 1993; Балановская и др., 1994в; Грехова и др.,1996]. Геногеографические карты главных компонент известны и для населения Западной, точнее, Зарубежной Европы [Menozzi et al., 1978; Piazza et al., 1981; Ammerman, Cavalli-Sforza, 1984]. Хотя наборы анализируемых генов для Западной и Восточной Европы не полностью совпадают, перекрывание наборов значительно, а результаты разительно отличаются и, можно сказать, географически диаметрально противоположны. В самом деле, значения 1-ой компоненты меняются в Западной Европе с юго-востока на северо-запад, то есть под прямым углом к направлению изменений этой же компоненты в Восточной Европе. Следовательно, для формирования западноевропейского генофонда главными оказались импульсы, шедшие не с востока, а с юга, точнее с юго-востока, с Балкан и Малой Азии на север и северо-запад вплоть до Британских островов. 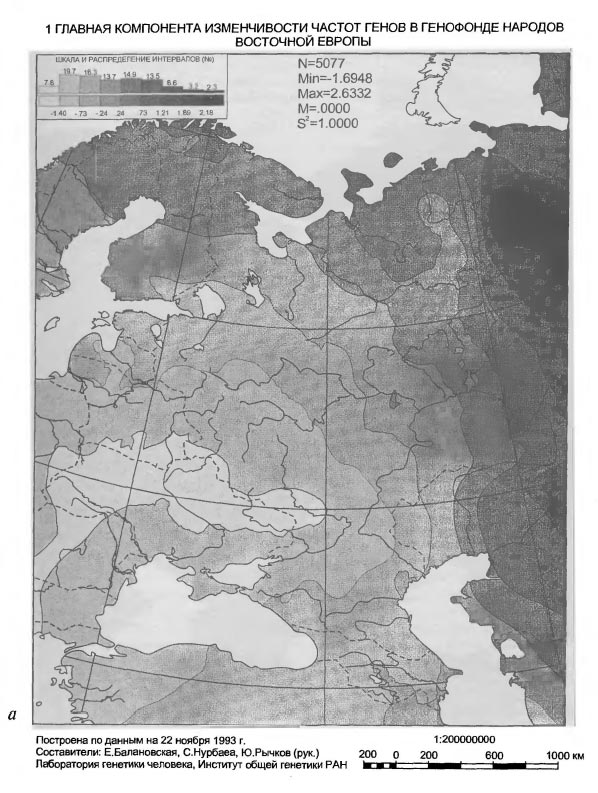
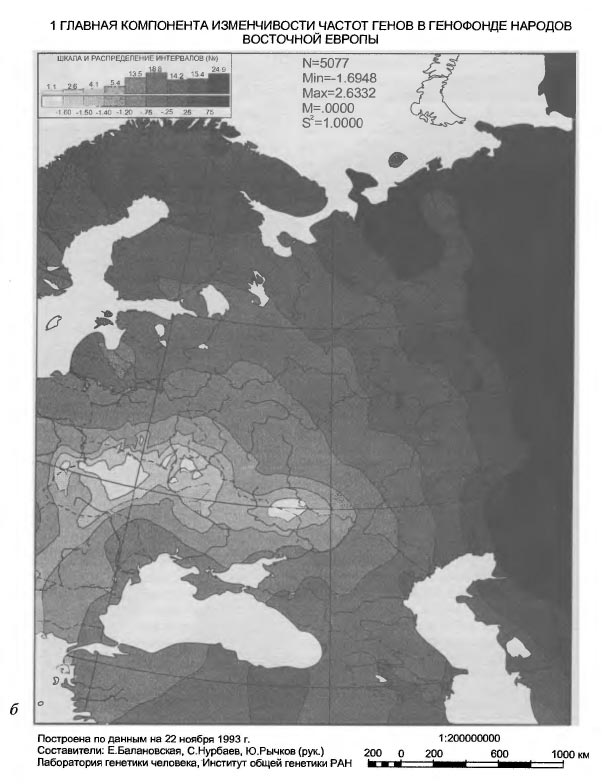
1 ГЛАВНАЯ КОМПОНЕНТА ИЗМЕНЧИВОСТИ ЧАСТОТ ГЕНОВ В ГЕНОФОНДЕ НАРОДОВ ВОСТОЧНОЙ ЕВРОПЫ Ниже мы увидим, что такое же (юго-восток - Северо-запад) географическое направление генетических импульсов имело место и в Восточной Европе, но здесь оно было в прямом смысле второстепенным, поскольку представлено на карте лишь 2-ой главной компоненты, по условию анализа имеющей меньший удельный вес в общей дисперсии генных частот. В свою очередь, для Западной Европы столь же второстепенными оказываются генетические влияния по линии Восток-Запад, они отражены лишь на карте 2-ой, а не 1-ой главной компоненты, как в Восточной Европе. Сравнительный картографический анализ двух генофондов позволяет уяснить главные факторы истории каждого из них. Современный западноевропейский генофонд приобрел основные черты своей географической структуры в эпоху так называемого неолитического перехода, связанного с зарождением и развитием земледельческого уклада жизни в Малой Азии и с проникновением через Балканы в Европу не только земледельческих традиций, но и волн земледельцев [Ammerman, Cavalli-Sforza, 1984]. Поэтому географическая структура тотального генофонда народов Западной Европы до сего времени коррелирует с изохронами радиоуглеродных датировок неолитической земледельческой культуры в Европе. Будучи ортогональны друг другу, западноевропейское и восточноевропейское направления изменений 1-ой главной компоненты обоих генофондов расходятся под прямым углом из общей вершины на Балканах. Поэтому карту 1-ой компоненты восточноевропейского генофонда можно было бы рассматривать по аналогии как след того же неолитического перехода. Действительно, при укрупнении генетического масштаба (рис.VI-3a) на карте, хотя и слабо, но все же видна связь клиновидной структуры юго-западного восточно-европейского минимума с Балканами и Пелопоннесом. Если еще более увеличить генетический масштаб (см. рис. VI-3б), можно даже обнаружить несколько микроядер - скоплений генов и генных комплексов - в пределах этого минимума. Некоторые из этих микроядер приходятся на ареал Трипольской культуры. И все же общую географическую структуру генофонда народов Восточной Европы, как она передается картой 1-ой компоненты, едва ли возможно связать с неолитической историей проникновения земледелия на этот субконтинент. Достаточно вспомнить, что распространение земледелия из области древней Трипольской культуры на северо-восток Русской равнины произошло на 3-4 тысячелетия позже времени существования этой культуры и связано не с движением демографической волны плотности неолитических земледельцев, как в Западной Европе, а с колонизацией Русской равнины историческими славянами [Седов, 1982; Рыбаков, 1993]. Рассуждение по аналогии с Западной Европой здесь не годится хотя бы потому, что имеет место смена географического направления 1-ой главной компоненты генофондов при переходе из Европы Западной в Восточную. Лишь западноевропейское направление юго-восток - северо-запад пока обосновано как направление движения волны земледельцев. Следует искать другие исторические причины особой географии восточноевропейского генофонда. Возможно, что не неолитический переход к земледелию, а пастушеское (транспортное и тягловое) скотоводство, коневодство и всадничество эпох энеолита, бронзы, а затем и раннего железа сыграли в Восточной Европе ту роль в создании географически упорядоченной структуры генофонда, которую неолитическая земледельческая революция выполнила в Западной Европе. Карта 1-ой главной компоненты, особенно в том, что касается юго-западной клинообразной структуры со множеством микроядерных скоплений генов в полосе от верховий Днестра до среднего течения Дона, вполне позволяет видеть в такой "ленточной" геногеографии лесостепной (и степной) полосы Восточной Европы генетические следы древних индоевропейцев энеолита и бронзы [Сафронов, 1989], а так же иранцев раннего железа, если к ним относить Геродотовых скифов-пахарей и земледельцев. Если же, вслед за Б.А.Рыбаковым [Рыбаков, 1993], видеть в сколотах восточную ветвь праславян, то тогда в карте 1-й компонеты восточноевропейского генофонда можно распознать и генетические следы славянской колонизации Восточной Европы. Четко выделяющаяся на карте 1-й компоненты юго-западная клинообразная структура приходится на лесостепную зону, наиболее благоприятную для обитания в самые разные исторические эпохи. Таким образом, не отдельная эпоха, культура или племя, а самые разные племена и в самые разные далеко отстоящие друг от друга времена - от палеолита до средневековья - могли оставить по себе память в генофонде современного населения, отпечатавшись в одной и той же части его ареала, так сказать, "след в след". Возможно, что именно географическая поясность природной среды, с оптимумом условий в лесостепном поясе Восточной Европы определила приуроченность к этому поясу и доминирование в разные эпохи одного и того же географического направления исторического процесса. Это направление, в силу устойчивой его повторяемости, в конечном счете оказалось решающим для судеб народонаселения Восточной Европы. Восточноевропейский генофонд приобрел устойчивую пространственную ориентировку, динамика генофонда стала географически закономерной. Эту доминирующую здесь закономерность - проникновение юго-западных, балканских, импульсов в пределы лесостепи с дальнейшей иррадиацией по Восточноевропейской равнине - передает ныне карта 1-й главной компоненты восточноевропейского генофонда. Таким образом, восточный и западный европейские генофонды складывались под давлением разных исторических обстоятельств, формировавших на Западе преимущественно земледельческие, а на Востоке - пастушеско-скотоводческие племена. Однако, оба направления исторического процесса, запечатленные в двух современных генофондах, берут начало все же в одной и той же области Европы - на Балканах. Эта, сохранившаяся и в современности, особая роль Балканского узла в истории Европы, как свидетельствует геногеография, определилась тысячелетия назад и определила собой историческую географию обоих европейских генофондов. Степь - Лес, 2-ая главная компонента изменчивости восточноевропейского генофонда 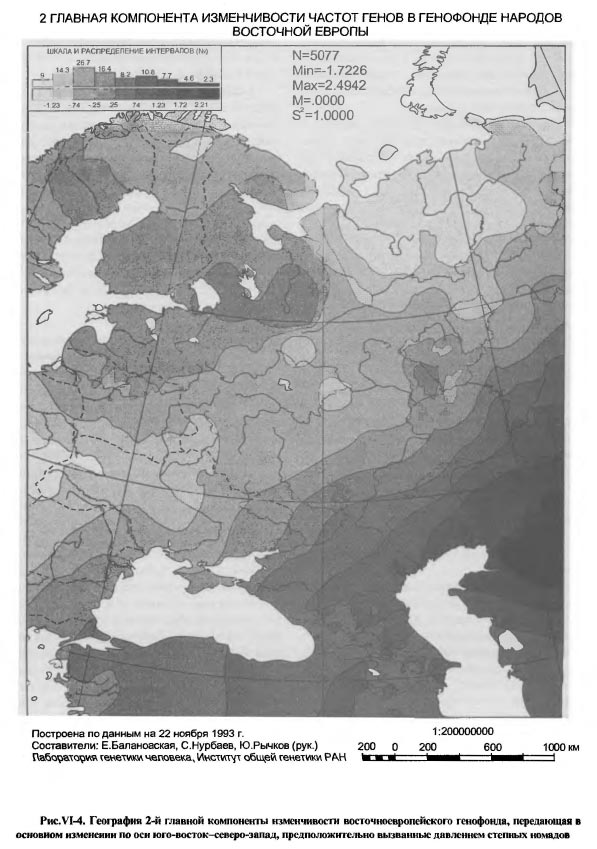
На карте 2-ой компоненты (рис. VI-4) подчеркнуты те изменения в восточноевропейском генофонде, которые идут в направлении юго-восток - северо-запад, от казахстанских, прикаспийских и северо-кавказских степей в Поволжье и в лесостепные пределы Русской равнины. Очевидна связь этого направления с имевшим место на протяжении нескольких тысячелетий давлением степных кочевников. Судя по тому, что на карте эпицентр этого давления в виде максимума значений 2-ой главной компоненты приходится на Приаралье, можно заключить, что особо значительное влияние на современную географию восточноевропейского генофонда оказали племена андроновской культурно-исторической общности, а затем скифо-сармато- аланские племена. Тюркское влияние на генофонд, хотя и трудно, но все же возможно вычленить из общей картины воздействия кочевого мира, если учесть, что 5-ый интервал шкалы карты, охватывающий Анатолию, оба побережья Боспора и Крым, наблюдается также в виде островка в низовьях р.Камы. Следы более ранних тюркских кочевников, по-видимому, полностью сливаются с общей географией воздействия номадов на восточноевропейский генофонд. Отметим, однако, что, судя по карте, с продвижением к Северу это воздействие довольно быстро затухало, не проникая севернее Волжской Болгарии. Так что даже монголо-татарское владычество не продвинуло границу степных генетических влияний к северу далее, чем она сложилась в древности. Выше уже отмечалось, что 2-я главная компонента восточноевропейского генофонда имеет такое же географическое направление, что и 1-я компонента западноевропейского генофонда. Но исторические причины, сформировавшие это единое направление генетической изменчивости населения на Западе и Востоке Европы совершенно различны, как различен и удельный вклад этих направлений в формирование структуры обоих генофондов: на Западе - это движение волны неолитического земледелия и земледельцев, начиная с V тыс. до н.э. и даже ранее, а на Востоке - повторяющиеся в течение нескольких тысячелетий, вплоть до позднего средневековья, волны степных номадов. На западе события эпохи неолита стали определяющими для всей последующей истории генофонда современного населения, а на востоке Европы непрекращающееся генетическое давление степных племен не распространилось севернее лесостепи и осталось второстепенным фактором сложения современного генофонда. Однако, фактор евразийских номадов не единственный, обнаруживающий себя в географии 2-й компоненты современного генофонда. На карте видны три другие полюса изменчивости: висло- волынско-днепровский, ладого-онежский и мезень-печерский, которые среди современных этнических групп легко сопоставляются: первый - с западными и восточными славянами, второй - с западными финнами, а третий - с восточными финнами и самодийцами. Все три как бы противостоят давлению степного мира, в результате чего в срединной части Восточной Европы возникает обширная "нейтральная полоса", своего рода генетическая Русская равнина. Ее пространственная ориентировка весьма напоминает культурно-историческую географию неолита и ранней бронзы на этой территории, такие общности как ямочно-гребенчатой и гребенчато-ямочной керамики, волосовскую, фатьяновскую [Крайнов, 1987а, б]. Висло-Волынско-Днепровское ядро на западе этой генетической равнины играет формообразующую роль, поскольку соответствует географии памятников шаровидных амфор и шнуровой керамики, находящихся в истоках фатьяновской культуры. Связана с этой генетической равниной и Южная Прибалтика - предполагаемая прото-фатьяновская территория [Крайнов, 19876, карта с.72]; Ладого-Онежское ядро вполне соответствует финской культуре ладьевидных топоров, ядро в Восточном Прионежье - позднекаргопольской культуре эпохи бронзы на Севере Восточноевропейской равнины [Ошибкина, 1987, карта с.148]; Мезень-Печерское ядро, занимающее обширную северовосточную область карты - неолитическим северным культурам ямочно-гребенчатой керамики, а в бронзовом веке - чойновтинской и лебяж- ской культурам [Ошибкина, 1987, карта с. 155]. Можно найти также указания на то, что современное Мезень-Печерское геногеографическое ядро (как и Карпатско-Днепровское ядро карты 1-й компоненты) уходит своими корнями в местный поздний палеолит [Балановская и др.,1994в; Грехова и др., 1996]. В целом же очевидно, что карта 2-й компоненты, как и 1-й, представляет синтез разных исторических эпох от палеолита, неолита и бронзы до позднего средневековья, Присутствие западного - Висло-Волынско- Днепровского - полюса изменчивости генофонда на картах не только 1-й, но и 2-й главной компоненты представляется неожиданным, поскольку противоречит принципу ортогональности (отсутствие общих элементов) главных компонент. Появление такого общего географического элемента в распределении обеих главных компонент изменчивости генофонда указывает лишь на то, что Карпато-Днепровская область Восточной Европы имела существенное значение во взаимодействии древних народов и культур не только в направлении Европа - Азия, но и в направлении Степь - Лес. Ключевая роль юго-западного региона Восточной Европы подтверждается тем, что этот регион сохраняет значение полюса развития восточноевропейского генофонда на картах не только первых двух, но и 3-й главной компоненты. Восточноевропейская равнина - скрещение исторических путей, 3-я главная компонента изменчивости восточноевропейского генофонда 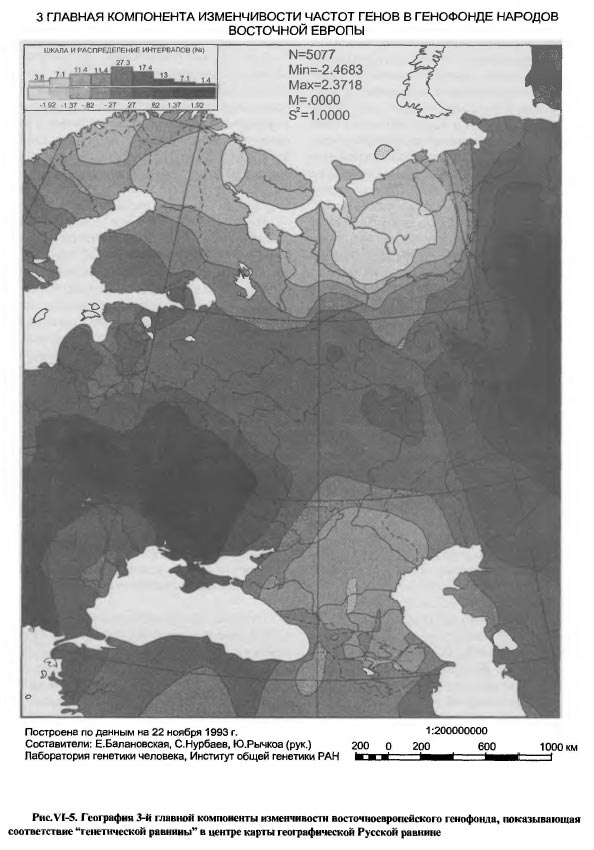
Отметим прежде всего отчетливость географической конструкции этой карты (рис.VI-6): два максимума с запада и востока и два минимума с севера (Кольско-Мезень-Печерский) и с юга (Северокавказский) простирают свои влияния на Восточноевропейскую равнину таким образом, что вся она превращается в обширнейшую генетическую равнину - область равнодействия противоположных влияний на генофонд. Этот же тип рельефа - равнина в окружении горных поднятий - был описан нами ранее по данным антропогеографии, картированным в виде отклонений от среднерусских величин ряда антропометрических и антропоскопических признаков [Рычков, Балановская, 1988]. Соответствие положения показываемой картой 3-ей компоненты генетической равнины физико-географическому положению Русской равнины явно неслучайно и подчеркивает географическую приуроченность исторического процесса. Генетическая однородность этого обширного пространства согласуется с фактами распространения на нем обширных и долговременных культурно-исторических общностей. Это неолитические культуры ямочно-гребенчатой керамики, неолит-энеолитическая волосовская общность, раннебронзовая фатьяновско-балановская общность, обширнейшая зона поселений-городищ эпохи железа [Крайнев, 1987а, б]. С фатьяновско-балановской общностью, по-видимому, связано явное соответствие Сура-Волжского ядра нашей карты исключительному скоплению в этом районе балановских древностей [Бадер, Халиков, 1987, карта 12, с.77]. Вся Русская равнина выглядит на этой карте как генетически однородное пространство, подготовленное историей для объединения его в последующие тысячелетия Северо-Восточной Русью, а затем и Московским государством. В целом, карты 3-й и 2-й компонент генофонда близки в том, что касается выделения Русской равнины как особой геногеографической структуры, но на карте 3-й компоненты контуры этой структуры ближе к действительным географическим. Близка к действительной и картина лесной зоны, передаваемая обеими картами. Принципиально новый элемент геногеографии, выявившийся именно на карте 3-й компоненты - это выделение Кавказа в один из полюсов развития восточноевропейского генофонда. Карта 3-й компоненты, как бы уточняя предыдущую, показывает, что степные влияния на Русскую равнину - это не только след номадов Приаралья, но и степняков Предкавказья, начиная, по крайней мере, с катакомбной культуры. Однако, как показывает карта, влияние кавказского фактора на формирование восточноевропейского генофонда не прослеживается на север далее Волго-Донского междуречья, то есть, оно невелико или же недостаточно постоянно. Два главных элемента пространственной структуры генофонда Таким образом, синтетические карты восточноевропейского генофонда выявили, в качестве главных элементов его историко-географической структуры, два наиболее постоянных. Один из них - это срединная область Восточной Европы - Русская равнина, она и в генетическом отношении представляет равнину - обширное пространство равновеликих значений. Здесь все идущие извне воздействия исторического процесса на генофонд взаимно нейтрализуются, и генетический рельеф нивелируется. Очертания этой срединной области отчетливо видны на картах 2-й и 3-й главных компонент, а карта 1-й компоненты показывает, что эта генетическая равнина представляет наклонную плоскость с поднятием от левобережного Приднепровья к Уралу. Такая география генофонда населения центральной части Русской равнины означает, что в нем весьма равномерно перемешаны и дисперсно перераспределены генетические влияния древних племен лесного Севера и степного Юга. Иными словами, встречные импульсы не столь сталкивались, сколь растекались по Русской равнине и нивелировались с течением исторического времени. Даже благоприятная география водных путей и широтная поясность природы Восточной Европы не помогли сформировать четкого широтного градиента в изменчивости генофонда. Градиент лишь намечен по южному и северному краям Русской равнины, указывая на то, что это направление исторического процесса не было преобладающим, хотя могло быть постоянным. Сложившийся в этих условиях генофонд населения срединной части Русской равнины, в принципе, соответствует представлению о среднеевропейском антропологическом типе. Однако геногеография указывает на то, что этот тип в Восточной Европе конвергентен, но не дивергентен по отношению к среднеевропейскому антропологическому типу Зарубежной Европы и имеет в Восточной Европе самостоятельное происхождение. Среднеевропейский антропологический тип в Восточной Европе имеет четкую отличительную особенность - выраженный градиент генетических изменений не столько с юга на север (от южно-европеоидного к северо-европеоидному), сколько с запада на восток, на уровне рас не второго, как в Западной Европе, а первого порядка. Другим устойчивым элементом историко-географической структуры восточноевропейского генофонда является его юго-западная часть в пределах от правобережья Припяти на севере до верховий Днестра и Днепровских Порогов на юге, от Закарпатья на западе до верховий Северского Донца, низовий Сейма и Десны на востоке. На картах разных главных компонент контуры этого генетического ядра несколько меняются, как бы передавая его географическую пульсацию в разные исторические эпохи. География этого ядра столь выразительна, что его связь с восточным славянством не требует дополнительных доказательств. Подчеркнем, однако, что силу генетической сути этого географического образования, его определение как восточнославянского, автоматически включает и все существовавшие на этой территории праславянские, дославянские - иранские и индоиранские - и доисторические палеопопуляции, чьи гены были унаследованы восточным славянством. Так, на карте 3-й компоненты это ядро, в целом, по географии повторяет ареал пражско-пеньковской культуры [Седов, 1982, карта 4], "сердцевина" ядра соответствует географии памятников Подольско-Днепровского лесостепного региона Черняховской культуры [Седов, 1982; Гей, 1993]. На карте 2-й главной компоненты видна связь этого же ядра с археологической областью племен лужицкой культуры 1-го тыс. до н.э. [Рыбаков, 1993, карты с. 15 и 27], а на карте 1-ой компоненты (при укрупненнии генетического масштаба) видно, что это генетическое ядро многосоставное, и расположение составляющих его элементов явно соответствует географии Геродотовых скифов-пахарей/сколотов (и гелонов), по реконструкции Б.А.Рыбакова [Рыбаков, 1979, карта стр.117; 1993; Мелюкова, 1989, карта 4, с.42-43] Если еще более углубиться в прошлое, то обнаружим соответствие локальных элементов этого ядра (в проекции 1-й главной компоненты) ареалам среднеднепровской, подкарпатской и других локальных вариантов культуры шнуровой керамики [Артеменко, 1987, карта 3, с.36), а общих контуров ядра - восточной части ареала культуры шаровидных амфор, свидетельствующей о появлении здесь древних индоевропейцев [Сафронов, 1989, карта рис. 45, с.351]. В еще большей древности это же многокомпонентное генетическое ядро соответствует общему ареалу неолитической трипольской культуры на разных стадиях и в разных локальных вариантах ее существования в правобережной и левобережной (софиевская группа) Украине. Как уже отмечалось выше, такую же географию, что и рассматриваемое здесь юго-западное генетическое ядро, имеет Вилендорф-Павлово-Костенковский позднепалеолитический культурный комплекс. Конечно, совпадение ареалов культур далекой древности с ареалом одной из географических структур современного генофонда не есть прямое доказательство генетической преемственности поколений на рассматриваемой территории, но является важным аргументом в пользу такой генетической непрерывности. Во всяком случае, это является основанием считать прикарпатско-среднеднепровский комплекс генов не только славянским, но унаследованным восточными славянами от индоевропейских (в том числе индоиранских и иранских) и более древних - неолитических - палеопопуляций лесостепной полосы Восточной Европы. Влияние этих палеопопуляций распространялось, как показывают наши карты, на всю Восточноевропейскую равнину от Причерноморья, Приазовья, Подонья и Поволжья до Прибалтики и Верхней Волги. Таким образом, геногеография, обнаружив приуроченное к лесостепи сильно структурированное скопление генов, указывает на то, что этот юго-западный Днепровско-Карпатский регион Восточной Европы имел краеугольное значение в формировании генетического фундамента восточноевропейского народонаселения и, особенно, восточных славян. В сущности этот геногеографический тезис вполне соответствует тому, какими виделись истоки основного антропологического типа восточных славян В.П.Алексееву [1969], который считал, что этот тип проникал с запада и юго-запада в пределы Украины, Белоруссии и Русской равнины. Именно этот путь проникновения, подчеркиваемого клинообразным ходом изолиний от Польши до Карпат, можно было видеть на карте 1-й компоненты географической изменчивости восточноевропейского генофонда (см. рис.VI-2), а на детализирующих вариантах этой карты (см. рис.VI-3 а, б), как и на карте 3-й компоненты (см. рис.VI-5) - уточнение юго-западной, балканской, ветви этого общего пути. Сказанное не следует понимать так, будто эти карты показывают географический след продвижения именно восточных славян в пределы Восточной Европы. Все сказанное на предыдущих страницах подчеркивало, что рельеф, показываемый картой - это кумулятивный отпечаток различных прошедших исторических эпох. Вполне исторически закономерно, что славянская колонизация Восточной Европы использовала эти проторенные тысячелетиями пути проникновения западных влияний. Вместе с тем, палеоантропологический анализ указывает на северно-южную дифференциацию ранних славян [Алексеева, 1973]. На представленных здесь картах главных компонент генофонда дифференциация этого типа видна лишь на карте 2-й компоненты (см. рис.VI-4), показывающей, однако, иные различия, чем между северными и южными европеоидами. Тем не менее, явные гено- географические следы участия северных европеоидов в формировании восточных славян будут представлены ниже, для этого потребуется изменение исторического масштаба карт генофонда. Подчеркнем еще раз, что из всех воздействш на этот генофонд за все время существования населения Восточной Европы главное формообразующее значение, упорядочившее пространст- венную структуру генофонда, имели воздействия в направлении запад-восток. Они носили характер проникновения с запада - юго-запада и распространения по Восточной Европе генных комплексов европейского и балканского происхождения и их носителей - древних народов и племен различных исторических и археологических эпох. Карта 1-й главной компоненты изменчивости генофонда показывает, как постепенно и достаточно равномерно население Восточной Европы генетически становилось европейским. В то же время, карта позволяет видеть, что эта генетическая "европеизация" отнюдь не дошла до конца. Возникло некое равновесие между европейскими и азиатскими влияниями, возникло своего рода евразийское равновесное состояние генофонда народов Восточной Европы. Это равновесие может быть лишь динамическим, а не стационарным, поскольку генофонд, как творение исторического процесса, может пребывать лишь в динамическом равновесии с состоянием окружающей исторической среды. Как совокупность генов, "вечных" в масштабе времени исторического процесса, генофонд передает нам информацию о наиболее устойчивых географических проявлениях этого процесса. Демонстрируемые географией главных компонент евразийские черты восточноевропейского генофонда и есть наиболее устойчивый генетический итог исторического процесса на территории Восточной Европы. Евразийская основа истории России состоит не в простом географическом факте ее одновременной принадлежности к Европе и к Азии, но и в том, что собственно европейская ее часть - Европейская Россия - генетически, а, значит, и исторически, формировалась как единство Запада и Востока, нерасчленимое на Европу и Азию, как историческая Евразия. Восточноевропейский генофонд в "русском масштабе" 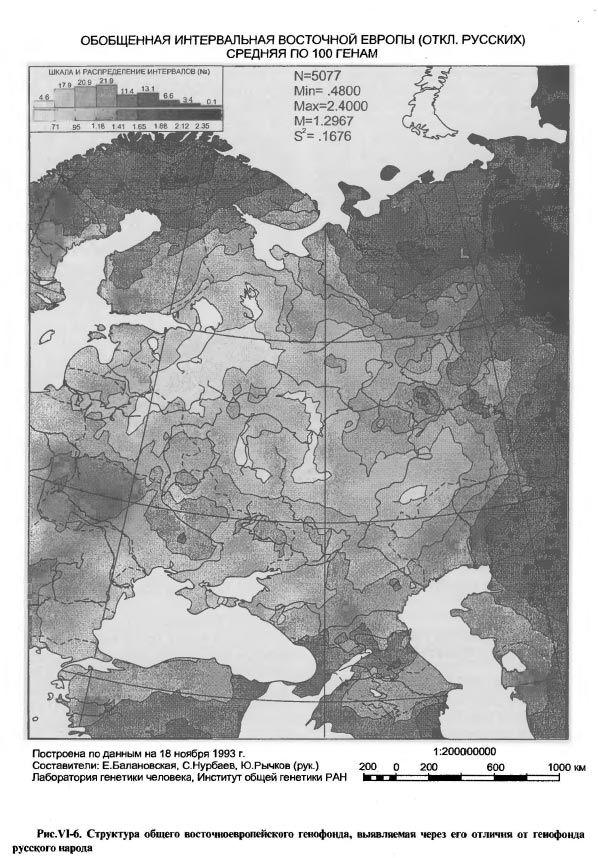
ОБОБЩЕННАЯ ИНТЕРВАЛЬНАЯ ВОСТОЧНОЙ ЕВРОПЫ (ОТКЛ. РУССКИХ) СРЕДНЯЯ ПО 100 ГЕНАМ 
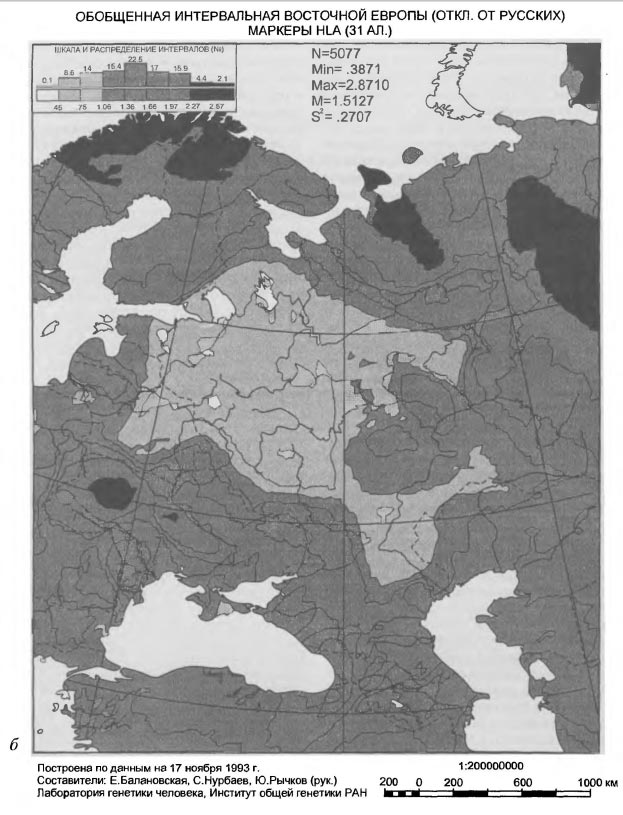
Карты предыдущего раздела изобразили срединную часть Русской равнины как генетическую равнину. Таким генофонд русского народа предстает в общем восточноевропейском масштабе: окружающие части общего восточноевропейского генофонда отличаются столь значительно, что в сравнении с этими различиями собственно русская часть выглядит пространственно огромной генетически однородной областью. Мы видим ее как бы на огромном удалении - на удалении большого исторического времени. Если же бросить взгляд из исторического ареала русского народа, мы должны увидеть те (и только те) структуры общего восточноевропейского генофонда, которые на протяжении последних одного-полутора тысячелетий имели значение для формирования генофонда русского народа. Переход к русскому этническому масштабу при рассмотрении особенностей общего восточноевропейского генофонда позволяет из всей суммы этих особенностей, возникших в самые разные исторические эпохи, выделить лишь те, которые имеют тот же исторический возраст, что и возраст русского народа. Коль скоро эта история достаточно молода, то особенности, относящиеся к русскому периоду общей многотысячелетней истории восточноевропейского генофонда, можно увидеть, лишь сменив исторический масштаб рассмотрения генофонда. На рисунке VI-6 весь восточноевропейский генофонд изображен в виде отклонений от средних частот генов русского народа и тех других народов, чьи ареалы полностью или частично входят в прямоугольное окно со сторонами 28° и 56° восточной долготы, 50° и 60° северной широты. В это окно попадает большая часть той генетической "равнины", которая обнаружилась ранее на картах главных компонент. Кроме русских, в него попадают частично северо-восточные украинцы, большая часть белоруссов, частично литовцы, латыши, эстонцы, вепсы, коми и коми-пермяки, полностью марийцы, удмурты, мордва, чуваши, татары, башкиры и отчасти даже казахи. Таким образом, лишь по территориальному преобладанию в этом окне русских, рассчитанные с компьютерных карт средние (взвешенные по площади занимаемой территории) генные частоты можно в первом приближении считать характеристиками русского генофонда (для более представительного анализа было бы необходимо учесть русских остальной территории Европейской России). Картированные отклонения от русского генофонда по 100 генам 34 локусов градуированы от минимума, означающего наибольшую близость к максимуму, указывающему на наибольшую удаленность от общерусских (средних) характеристик генофонда. Поэтому естественно, что на карте основной ареал русского генофонда выделился как светлая область (вторая градация шкалы), простирающаяся с северо-запада на юго-восток и с юго-запада на северо-восток. Такое простирание легко объяснимо из рассмотренной перед этим географии первых двух главных компонент восточноевропейского генофонда. В контурах ареала отражены основные моменты формирования русского генофонда в ходе славянской колонизации Русской равнины - движение на северо-восток, а также в противостоянии давлению степных кочевников с юго-востока и скандинавско- балтскому давлению с северо-запада. В целом же, по географическому размещению светлых и темных тонов на карте Восточной Европы, русский генофонд выглядит как часть общеевропейского, углубившаяся на восток в Заволжье, Приуралье и Прикаспий. По всему фронту этого движения виден резкий перепад величин отклонения от малых к большим, своего рода подножие крутого подъема, за которым далее лежит обширнейшая область высоких (явно не русских) значений - плато азиатского генофонда. Напротив, в исходном направлении на запад-юго-запад до самого края картируемой территории, до Польши, Чехословакии, Венгрии, Балкан, Босфора и Дарданелл, простирается область относительно малых - ниже средних и средних - отличий от русского генофонда, что еще раз и по-новому свидетельствует о продвижении европейских генных комплексов с Балкан и из Центральной Европы в Восточную Европу с ходом ее славянской колонизации и далее в пределы русского этнического ареала. Вглядевшись в этот высветленный на карте ареал, обнаружим два уровня близости всего восточноевропейского генофонда к общерусским характеристикам, один - более общий (по шкале значений он - второй), охватывающий северо- запад, центр и юго-восток России, и другой - уровень наибольшей близости (минимальный интервал шкалы карты), приходящийся, главным образом, на Новгородскую, Смоленскую и Тверскую области и, меньшими островками, на земли Владимирщины, Рязанщины, Орловщины и Оренбуржья. Оренбургский минимум является, скорее всего, исторически недавним следом развития Российского государства, отголоском какого-то направленного колонизационного потока. В других же названных минимумах угадываются генетические следы летописных новгородских словен, полоцких, смоленских и тверских кривичей, а также других славян времен существования Черниговского, Владимиро-Суздальского и Рязанского княжеств. О возможности такого прочтения геногео-графической карты говорит и территориально небольшая область повышенных (удаленных от русских) значений в верховьях р. Москвы, указывающая на генетические следы летописной голяди. В целом площади областей минимумов - наиболее "русских" значений генофонда - убывают с северо-запада на юго-восток так, что в их географическом изменении угадывается движение исторического времени развития русского народа при все большем включении в его генофонд местных, до-славянских, генных комплексов. Таким образом, ядро русского генофонда находится на северо-западе русского этнического ареала и включает часть белорусского ареала, но нигде не заходит в пределы современных ареалов народов Прибалтики. Отличие от балтов видно на карте довольно отчетливо. Если считать это ядро связанным с теми североевропейскими антропологическими элементами в составе русских (и белоруссов), которые генетически не улавливались анализом главных компонент в большом общем размахе разнообразия восточноевропейского генофонда, то, следуя выводам палеоантропологи ческого анализа [Алексеева, 1973], в этом же русском геногео- графическом масштабе должен обнаружиться среди восточных славян и комплекс генов южного - средиземноморского - происхождения. Как показывает карта (см. pиc.VI-6), этот комплекс, действительно, обнаруживается, он локализован точно в пределах Украины, будучи подразделен на западную и восточную (поднепровскую) части. Принцип картирования отклонений от общих свойств русского или какого-либо иного генофонда может быть применен не только ко всей анализируемой совокупности генов, но и к отдельным генным комплексам, отражающим разные этапы истории народа и его генофонда. На рисунке VI-7 показаны результаты такого картирования отклонений от свойственных русским частот двух групп генов. Для генов, определяемых через полиморфизм различных белков крови (см. рис.VI-7а), карта передает достаточно древний этап славянской колонизации Русской равнины, с отчетливым сохранением генетической близости восточных славян с южными. На карте она видна как сплошной меридионально протянувшийся пояс малых (то есть, "русских") значений от Ладоги и Онеги до южных границ Болгарии и особенно малых значений на Валдае, в срединной части Белоруссии и в Карпатах. Если этот пояс - не след продвижения славян на север Восточной Европы, то весьма вероятно, что это генетический след событий VI века, характеризуемых историками как "возвратное движение на покинутый некогда юг" и "массовое вторжение славян на Балканский полуостров" [Рыбаков, 1993, с.47, 48]. При таком прочтении наша генетическая карта показывает такое же дублирование на Балканах "русских" генетических характеристик, какое отмечается историками в отношении имен славянских племен, пришедших с севера. [Рыбаков, 1993, с.49]. Наиболее выразительной является карта (см. рис. V1-7б) отклонений от общерусского генофонда по генам, контролирующим комплекс лейкоцитарных антигенов человека (HLA), известный своей высокой дифференцирующей способностью и приуроченностью различных вариантов этого комплекса к различным этническим группам. На этой карте ареал русского генофонда предстает и сегодня разделенным на западную и восточную части по 42° в.д., по меридиану Ярославля, Суздаля, Владимира и Рязани, генетически воспроизводя в современности восточную границу древнерусского государства IX-XI века. Воспроизведение этой границы на генетической карте было в свое время объяснено на материалах антропогеографии русского народа, в которых эта граница проявилась с такой же отчетливостью [Рычков, Балановская, 1988]. Объяснение сводилось к тому, что до достижения этой восточной долготы славянская колонизация представляла собственно продвижение летописных славян на восток и их расселение по все большей территории, что должно было сопровождаться уменьшением плотности славянского населения. Генетическая и антропологическая граница возникла у того нижнего предела этой плотности, за которым колонизационный процесс или должен был иссякнуть, или мог продолжиться, перейдя в фазу культурной ассимиляции местного дославянского населения с помощью городов и монастырей как центров христианизации и славянизации окружающих эти форпосты финно-угорских племен, на что указывают и археологи [Седов, 1982]. Видя на геногеографической карте еще более четкое воспроизведение этой границы, чем на карте антропологической, мы можем лишь подтвердить предложенное ранее объяснение. Восточнее этой границы русский генофонд несет значительную долю генных комплексов до-славянского, главным образом финно-угорского, происхождения, включенных в русский генофонд после того, как в ходе колонизации процесс расселения сменился процессом культурной ассимиляции. В силу большей этнодифференцирующей способности множественных генов HLA, исходное ядро русского генофонда на карте (см. рис.VI-7б) отчетливо противопоставляется ядру украинского генофонда, однако, причины столь крайних различий (субминимум и субмаксимум шкалы), явно находящихся у пределов восточнославянской общности, но связанных лишь с определенным комплексом генов, требуют дополнительного генетического анализа, выходящего за рамки работы. Коль скоро оба полярных генофонда - русско- белорусский на севере и украинский на юге - оказались, действительно, у генетических пределов восточнославянской общности, упрощенно можно соотнести эти пределы с венедской и антской ветвями раннесредневекового славянства. * * * Заканчивая очерк исторической геногеографии Восточной Европы, подчеркнем, что он полностью основан на сопоставлении геногеографических карт с картами археологическими. В поиске соответствий между этими типами карт авторы исходили из соображения о том, что, если когда-либо в прошлом существовали условия, способствовавшие обособлению, сосредоточению и воспроизведению в ряду поколений определенной материальной культуры на определенной территории в определенный период исторического времени, то этих условий достаточно, чтобы в ареале этой культуры сложились отличительные особенности генофонда населения, создавшего эту культуру (или принесшего ее с собой). Поэтому, если археолог имел основания выделить культуру и наметить ее ареал на карте, то этого достаточно для сопоставления археологической и генетических карт, но не гарантирует обнаружения соответствия между ними. Чтобы такое соответствие обнаружилось, нужно, чтобы специфика генофонда, сложившегося в выделенном археологом ареале, была донесена генами из древности в современность. За прошедшее время от населения, некогда создавшего материальную культуру, может не остаться никакой иной памяти, кроме генов, перешедших по цепи поколений из древнего генофонда в современный и сменивших на этом пути многих носителей с их этнонимами и культурной спецификой. Важно только то, чтобы эта цепь ни разу не прервалась, и тогда в современности может возникнуть генетический след древней археологической культуры в виде определенного генетического ареала. Именно этот ареал и требуется сопоставить с ареалами археологических культур. В работе представлены фрагменты таких сопоставлений, их следует рассматривать лишь как первую попытку такого рода, скорее показывающую возможность и некоторые правила перевода геногеографических карт на язык археологии, без претензии на точность перевода, поскольку это требует профессиональных археологических знаний и знания археологических источников. Археолог, если заинтересуется показанной здесь возможностью увидеть в географии генов современного населения прямое наследие прошлого разных археологических эпох, предложит заведомо более содержательное прочтение геногеографических карт. Сосредоточившись на их исторической интерпретации, авторы сознательно оставили в стороне экологическую интерпретацию карт, которая требует отдельного обоснования и исследования. Настоящая работа выполнена при частичной финансовой поддержке Государственных научно-технических программ "Приоритетные направления генетики" и "Биологическое разнообразие". |
загрузка...