4. Объективность геногеографии
|
Суха теория, мой друг
Но древо жизни вечно зеленеет Гёте. «Фауст» В предыдущих разделах мы бегло осмотрели понятийный аппарат и арсенал геногеографии. По ходу книги по мере необходимости давались «технические» пояснения. Но чтобы увидеть единство этой книги, анализирующей разные признаки, по-разному рассказывающие о русском генофонде, надо ответить на ряд важнейших общих вопросов. Без ответа на них результаты останутся отдельными страницами разрозненных томов. Например, насколько изложенные выше положения о структуре генофонда являются спорными? Получим ли мы с помощью арсенала геногеографии действительно объективный портрет генофонда? Как глубока генетическая память? Насколько далеко в глубь времен проникают генетические «зонды»? Соответствуют ли данные генетики данным других наук - археологии, истории, демографии, антропологии, лингвистики? Насколько надёжны и устойчивы оценки изменчивости генофонда? Можно ли в данных о совершенно разных признаках разглядеть общую картину изменчивости генофонда? В этом разделе мы попробуем дать краткие ответы на эти первостепенные вопросы. Поговорим, например, о важной характеристике генофонда - его разнообразии (дифференциации). Выше мы утверждали, что если возьмем средний по множеству генов показатель различий между популяциями FST, то получим такую интегральную оценку дифференциации генофонда, которая уже не зависит от неравной приспособленности аллелей к среде и потому называется «селективно-нейтральной». Иными словами - не зависит от действия отбора. Эта оценка определяется только историей генофонда (FST = Fe) и зависит только демографического облика популяции: от соотношения исторически сложившихся размера популяций Ne и миграций Me: FST≈Fe=1/(4NeMe+1). Это положение - одно из самых важных для понимания изменчивости генофонда и истории его сложения, как бы мы его ни изучали: с помощью многомерной статистики или компьютерных карт. Однако именно оно зачастую вызывает сомнения. Действительно, казалось бы, как можно по генам, каждый из которых потенциально подвержен отбору, получить портрет генофонда, сформированный только историей? Получить селективно-нейтральный портрет генофонда, не зависящий от отбора, не зависящий от биологической функции гена, не зависящий от среды? Можно ли найти доказательства этому странному положению? Чтобы дать убедительный ответ на этот один из самых коварных вопросов - а без ответа на него нет смысла описывать ни русский, ни любой другой генофонд! - мы приведём два ряда доказательств: §1 и §2 данного раздела. Эти доказательства выходят далеко за пределы не только русского генофонда, но и Европы. Но лишь охватив изменчивость многих генофондов, в больших масштабах времени и пространства, можно проверить основную гипотезу: гипотезу выполнения равенства FST≈Fe=1/(4NeMe+1).. Первый ряд доказательств относится к генохронологии (§1). Генохронология по данным генетики оценивает возможное время реальных исторических событий. Она дает датировки давно прошедших событий в истории генофонда. Причём _все её датировки основаны на предположении равенства FST≈Fe=1/(4NeMe+1).. Поэтому генохронология дает одну из самых очевидных возможностей проверки этого равенства. Если генетическая датировка будет соответствовать исторической дате события, то гипотеза FST≈Fe верна, и мы действительно можем реконструировать селективно-нейтральную историю генофонда по совокупности генов, подверженных отбору. Второй ряд доказательств (§2) относится к прямому сопоставлению оценок разнообразия генофонда, полученных по данным генетики (FST) и по данным других наук (Fe). При этом для одного и того же генофонда мы получаем две независимые оценки его дифференциации. С одной стороны, оценку дифференциации генофонда FST, полученные по множеству генов. А с другой стороны - оценку дифференциации Fe, полученную по информации, вообще никакого отношения к генам не имеющей: по чисто демографическим оценкам размера популяций Ne и миграций Мe; или же по данным о распределении фамилий. Если такие ряды независимых оценок будут соответствовать друг другу - то мы получим веское доказательство справедливости основной гипотезы FST≈Fe=1/(4NeMe+1)). Оба ряда приводимых доказательств созданы благодаря таланту и усилиям Юрия Григорьевича Рычкова и его школы. Они сводят воедино данные многих исследователей, многих научных работ по изучению генофонда народов СССР, которые специально планировались профессором Ю. Г. Рычковым для проверки равенства FST≈Fe и были проведены под его руководством. Эти данные публиковались в целом ряде статей в разном составе авторов, однако сейчас незаслуженно забыты. Мы приведём эти результаты так, как они изложены в наших совместных публикациях с Ю. Г. Рычковым: описание генохронологии (§1) приводится по книге Е. В. Балановская, Ю. Г. Рычков «Геногеография (гены человека на карте СССР)» [Балановская, Рычков, 1990а]; сопоставление генетических и не генетических оценок дифференциации (§2) - по статьям цикла Е. В. Балановской и Ю. Г. Рычкова «Этническая генетика» [Балановская, Рычков, 1990б,в; Рычков, Балановская, 1990а]. Мы старались - вопреки вынужденным сокращениям - полностью сохранить дух и стиль изложения этих работ. Хотя обобщение и описание этих результатов приведено в наших общих с проф. Ю. Г. Рычковым публикациях, однако основная часть данных была получена задолго до начала нашей совместной работы. Поэтому, не беря на себя смелость редактировать или исключать те черты, которые кажутся нам отнюдь не бесспорными, из уважения к памяти Ю. Г. Рычкова мы приводим их именно в том виде, который его удовлетворял. В §3 мы рассмотрим, от чего зависит устойчивость оценок изменчивости генофонда, в §4 дадим обзор генетической изменчивости в основных регионах мира, а в §5 немного поговорим об истории нашей науки. §1. ГенохронологияМЕРА - ПОКОЛЕНИЕ Человеческие гены вовлекаются в исторический процесс, поскольку человек не только творец истории и исторического времени, но и творение этой истории. Это значит, что генетические процессы, происходящие в человечестве, не только регулируются историей, но и происходят в историческом времени. Причём гены обеспечивают людям всего лишь возможность кратковременной жизни, тогда как люди обеспечивают генам возможность передачи по длинной цепи поколений. Поколение - естественная генетическая единица времени. А смена поколений - это ход часов, отмеряющих время течения генетических процессов. Но поколение - это ещё и определённый период в социальной истории человека. Поэтому можно два ряда событий (естественной истории населения и его социальной истории) выразить через одну единицу времени - поколение. Историческая наука при датировке пользуется иным - астрономическим временем, к которому сводимы любые календарные системы - линейные и циклические. Есть ли здесь место поколению? Можно ли представить в поколениях хронологию всеобщего или локального исторического процесса, не возвращаясь при этом к «доисторическому» способу времяисчисления, когда заучивание каждым своего места в генеалогической цепи, тянущейся от предков-основателей, было равносильно изучению истории и познанию исторического времени? Термин «историческое событие» в его прямом значении - событие в истории. Но обычно историческим событием мы называем то, что (как мы сейчас думаем) не исчезнет из истории в будущем, то, что войдёт в нее. Генохронология датирует такие события, которые и в прямом, и в переносном смысле являются историческими: только оставив свой след в истории дальнейших поколений, эти события могут быть уловлены генетическим методом. ИСТОРИЧЕСКОЕ СОБЫТИЕ Будем считать генетически значимым такое историческое событие, которое привело к изменению дифференциации генофонда [Fe = 1/(4NeMe+1)], то есть затронуло важнейшие параметры генофонда - дрейф генов Ne и миграции генов Me, рассмотренные нами выше (Приложение, раздел 1, §3). Например, это может быть изменение типа хозяйства или даже какой-нибудь конкретной технологии культуры, которое привело к росту эффективности хозяйства. А затем через это изменение - к приросту численности населения, к изменению возрастной структуры, а значит, и к изменению генетического размера популяции Ne. Но цепь генетических последствий такого события на этом может не оборваться и продолжиться дальше в форме увеличения подвижности населения, роста культурно-экономических контактов и так далее, что непременно скажется на изменении показателя миграции Мe. Таких событий - множество. Намного сложнее найти пример такого события общественной истории, для которого можно было бы категорически отрицать его генетические последствия. Вся разница лишь в длине цепи и отдалённости генетических последствий от их исторической первопричины. Так, события в области не только материальной, но и духовной культуры потенциально являются и событиями в истории генофонда. Наглядными примерами могут служить широко известные генетические последствия событий в истории мировых религий и малых сект (генетикам хорошо известно чрезвычайное генетическое своеобразие ряда сектантских общин, заключающих браки только с единоверцами). Для формирования русского генофонда важнейшее значение имели духовная мощь русского монашества и его распространение за пределы этнического ареала. Монастыри и одинокие скиты не приносили гены на окраины ареала, где они основывались в надежде уйти от суеты мирской жизни. Но они становились форпостами русской культуры, центрами духовной и культурной жизни, вокруг которых организовывалась общественная и экономическая жизнь (вспомним хотя бы, что важнейшие ярмарки России проводились у монастырей, куда съезжался православный люд), возникали новые крупные популяции. ГЕНЕТИЧЕСКИЕ ЧАСЫ Современная наука предлагает немалый набор самых совершенных часов для измерения времени в различных эволюционных процессах. Но для анализа генетических последствий они не годятся. Ведь радиоуглеродные часы отсчитывать время, прошедшее с момента гибели какого-либо существа, будь то водоросль, дерево, бабочка или человек. Они имеют отношение к хронометрированию именно той истории, которая когда-то (и часы должны указать, когда именно) оборвалась. Здесь все зависит от того, успел ли тот человек, кости которого из археологических раскопок попали в радиоуглеродную лабораторию (или в лабораторию палеоДНК), оставить потомков или передать какую-либо иную весть в будущее. И если нет, то такие часы будут указывать всего лишь время обрыва какой-либо цепи. Получается, что, располагая совершенным хронометром, мы не имеем никакой гарантии и можем лишь надеяться, что датируемое им событие прошлого имело отношение ко всему последовавшему за ним будущему. Парадокс фантастической бабочки Рэя Брэдбери, описанный одновременно с изобретением радиоуглеродных часов, имеет прямое отношение к ним. Поэтому воспользуемся поколением как единицей времени. Основываясь на исследованиях, проведённых генетиками и антропологами, можно считать, что у человека, к какому бы кругу народов и культур он ни принадлежал, средняя длительность поколения составляет около 25-26 лет, если за точку отсчёта взять средний возраст родителей при появлении у них первого ребенка. В таком темпе происходит нарождение новых поколений у папуасов Новой Гвинеи и москвичей, у пигмеев Конго и армян, у эскимосов и чукчей Чукотки и литовцев, у алеутов Командорских островов и уйгуров Средней Азии, у удэгейцев дальневосточного Приморья и монголов. Итак, используя гены современного человека как часы, отсчитывающие время генофонда, мы точно знаем, что (в отличие от калий-аргоновых и радиоуглеродных часов) у этих генов были свои предшественники в каждом предыдущем поколении, и что время, отсчитываемое поколениями, не прерывалось ни разу. Точка отсчёта времени всегда условна - ведь и сегодня на традиционных календарях разных народов одни и те же события приходятся на разные даты из-за разных точек, выбранных в качестве начала летоисчисления Поэтому мы примем за начальный момент для обратного отсчёта времени год генетического изучения тех популяций, события в истории которых мы попытаемся отыскать в генетической летописи. ДАТИРУЕМЫЕ СОБЫТИЯ Генохронологически датированные события расположим в хронологическом порядке: от совсем недавних времен - к древним2. ХОТОНЫ. На крайнем северо-западе Монголии проживает небольшая этническая группа - хотоны. Как этнос они начали формироваться около 250 лет назад, когда во время войн джунгаров пленные (из узбеков, казахов, уйгуров и других народов Центральной Азии) были поселены в этом уголке Монголии в надежде, что они наладят в этой скотоводческой стране земледелие. Хотя навыки к земледелию, принесенные с родины, в новой окружающей этнической среде постепенно утрачивались, но новая этническая общность - хотоны - все же возникла. После революции в Монголии хозяйство хотонов претерпело резкую реорганизацию. Их земледелие оказалось столь малоэффективным, что в 1934 г. специальным постановлением хотоны были переориентированы на новый для них тип хозяйства - скотоводство. Это постановление повлекло полную реорганизацию и всей популяционной структуры хотонов, в том числе структуры брачных связей, а значит, и миграций генов. Как видим, цепь, связывающая исторические события с возможными генетическими последствиями, здесь очень коротка. Запомним дату события: 1934 год. Генетически хотоны были исследованы под руководством Ю. Г. Рычкова монголом Ж. Батсуурем в 1978 г. Эта дата будет точкой обратного отсчёта генетического времени. УЙГУРЫ и ДУНГАНЕ. В связи с историей хотонов мы уже упомянули уйгуров - древний народ Центральной Азии с весьма трудной исторической судьбой. Была эпоха их гегемонии в этом районе мира, расцвет культуры, собственная письменность, но их силы истощились в борьбе с пришельцами, завоевавшими их родину. Один из актов этнической трагедии происходит в 70х годах XIX века, когда в борьбе за самосохранение уйгуры примкнули к восстанию дунган. Дунгане - ещё одно многомиллионное национальное меньшинство Китая, также испытывавшее притеснения. Дунгане и уйгуры в этом восстании потерпели сокрушительное поражение от регулярной китайской армии. В поисках спасения уйгуры и дунгане отдельными группами и семьями устремились из разных районов через горные перевалы в среднеазиатские пределы России. Русским пограничникам было дано указание, пропустив беглецов, закрыть границу перед преследовавшими частями китайской армии. Эти события происходили с осени 1870 по 1881 г., когда граница была окончательно закрыта. Но до 1884 г. мелкие группы беженцев просачивались через границу. В Средней Азии им были отведены территории для поселения. Часть уйгурских и дунганских семей поселилась в Ферганской долине, где 100 лет спустя, в 1970 г., их современные поколения, образовавшие несколько популяций, были изучены под руководством Ю. Г. Рычкова уйгуром Х.А. Каримовым. Итак, датировка описанных событий и начала формирования в Фергане популяций уйгур и дунган: 1870-1884 гг. Точка обратного отсчёта времени: 1970 г. АЛЕУТЫ. Из конца XIX века переместимся в его начало. Тогда в число территориальных владений России входила Русская Америка, простиравшаяся от Аляски на севере до Калифорнии на юге. Географически и экономически к ней относились и Алеутские острова, хотя административно они управлялись не из Новоархангельска - столицы Русской Америки, а из Охотска, игравшего тогда роль форпоста России на Тихом океане, которая позднее перешла к Владивостоку. Алеутские острова издревле, как указывают радиоуглеродные датировки археологических и палеоантропологических материалов, на протяжении 8 тысяч лет, были заселены алеутами - народом, родственным эскимосам, но с иной специализацией традиционного хозяйства. Что же касается Командорских островов, лежащих между Алеутской грядой и Камчаткой, то они не были обитаемы ни в древности (по данным археологической разведки), ни в момент их открытия при кораблекрушении экспедиции Беринга в 1741 г., ни позднее вплоть до 1826-1829 гг. (если не считать временно высаживавшихся там промысловых артелей русских, добывавших песца и морского зверя и возвращавшихся с добычей на Камчатку). В документах Российско-Американской компании сохранилось одно яркое свидетельство необитаемости островов в начале XIX в. С одного из кораблей Российско-Американской компании сообщалось (командиром брига «Финляндия» штурманом И. Ф. Васильевым), что в 1812 г. с острова Беринга им был снят один русский, оставленный там своей артелью на год охранять добытые шкуры, которые не могли увезти с собой, да забытый на целых три года. Другие его семеро товарищей, в свою очередь, были высажены в 1805 г. на острове Медном и забыты там на целых семь лет [Русская Америка, 1979]. Так, эта русская робинзонада определённо указывает, что ив 1812 г. Командоры все ещё были необитаемы. Наконец, было решено основать на Командорах постоянное поселение, которое служило бы и стоянкой для судов — единственной связи Русской Америки с Россией. Но поскольку Русской Америке и так остро недоставало русских, на Командорские острова было решено переселить алеутов с Алеутского архипелага. Осуществлял этот проект Ф. П. Литке во время кругосветной экспедиции на военном шлюпе «Сенявин» в 1826-1829 гг. Этими годами (1826-1829 гг.) и датируются два исторических события: Командорские острова утратили очарование необитаемости, но в составе народов России появился новый народ - алеуты (Алеутские острова ныне относятся к США). Их генетическое исследование было проведено Ю. Г. Рычковым вместе с И. В. Перевозчиковым и В. А. Шереметьевой в 1970 г. ВОСТОЧНО-САЯНСКИЕ ТЮРКИ. Спустимся далее в глубь времен, пропустив ещё тысячу триста лет. Следующая группа народов - тюркоязычные тувинцы, тоджинцы и тофалары в Саянах. Напомним, что ныне тюркские языки распространены от Колымы на северо-востоке Евразии до Босфора на юго-западе Евразии. Но предполагаемая их родина - в Алтае-Саянском нагорье и прилегающих районах Монголии. Время появления тюрок на исторической арене зафиксировано в древнекитайских летописях. Древний Китай на протяжении тысячелетий вел с переменным успехом, но в целом наступательные войны за овладение Центральной Азией, поэтому естественно, что все происходящее в этом регионе, тем более сложение нового народа -потенциального противника - привлекало пристальное внимание и фиксировалось в хрониках. Первое упоминание о тюрках относится к 518 г., и в нем уже встречаются этнонимы «теле» и «тубо», которые и сегодня звучат в названиях народов: «теле» - на Алтае (теленгиты, телеуты, телесы), «тубо» - на Алтае и в Саянах (тубалары. тофалары, тувинцы). В период 626-659 гг. погибли две тюркские империи — Западный Каганат и Восточный Каганат, между которыми Саяны составляли северный пограничный район. Выход из-под центральной власти дал начало самостоятельному развитию тюркоязычных народностей на Северной (Сибирской) окраине Внутренней Азии. Можно предположить, что это произошло в названный период распада империи: 626-659 гг. А самая ранняя дата из истории тюрок - 518 г. также может служить вехой в генетическом путешествии в прошлое. ТОФАЛАРЫ и ТУВИНЦЫ. Другими событиями с зафиксированными датами история Саян небогата. Но, пожалуй, можно попытаться отыскать в генетике современных популяций след ещё одного события. В конце жизни Чингисхана (умер в 1227 г.) монголы покорили саянских тюрок. Владычество монголов в Саянах длилось с XIII по XVI вв. Еще по указанию Чингисхана были созданы золотые прииски в самом центре земли тофаларов. Прииски действовали и века спустя, пока на огромной площади не были сведены все леса, пошедшие на костры для разогрева мерзлой земли в штольнях. И ныне этот район - на протяжении дня пути через него - полностью лишен леса и обходится тофаларами, чье традиционное хозяйство базируется на охоте и транспортном оленеводстве. Так возникла экологическая граница, разъединившая популяции тофаларов, живущих по разные стороны зоны золотых приисков. Поэтому попытаемся с помощью генохронологии отыскать в генах современных тофаларов след событий, связанных с монгольским завоеванием Саян и происходивших в XIII-XVI вв. Генетическое исследование тофаларов и других групп саянских тюрок было проведено Ю. Г. Рычковым со студентами Московского университета в 1964 г. ТУНГУСО-МАНЬЧЖУРЫ ПРИМОРЬЯ. Углубимся в прошлое ещё на тысячелетие и переместимся на Дальний Восток, в Приморье, где ныне проживают тунгусо-маньчжурские народности: удэгейцы и нанайцы, по традиционному укладу -рыболовы и охотники. Теперь их всего несколько сот человек в двух посёлках. Однако за той картиной демографии и быта, которую застали русские в момент присоединения Приморья к России, скрывалась длительная и насыщенная событиями история народов Дальнего Востока. Она восстановлена российскими исследователями и проконтролирована по все тем же древнекитайским хроникам. Некогда население этого края было столь многочисленным, что в Приморье и соседней Маньчжурии возникали мощные племенные союзы и государства с городами, укреплениями, регулярным войском и высокой культурой, соперничавшей порой с древнекитайской. Знаменитый Конфуций посетил этот край. Первые отрывочные сведения об этих племенах восходят ещё ко II тысячелетию до н.э., а с середины I тысячелетия до н.э. они становятся уже постоянными в китайских хрониках. Еще позднее эти племена становятся известны как создатели Золотой империи чжурчженей, затем государств Мохэ и Бохай. В конечном счете, все они потерпели поражение в борьбе с Древним Китаем, а затем и с монголами. У современных потомков этих блестящих предшественников на момент их генетического обследования все ещё сохранялись элементы преемственности материальной и духовной культуры. Так, один из мифов, сохранившихся до наших дней, описывает, как однажды на небе вспыхнуло сразу десять солнц, испепелив на Земле все живое и расплавив Землю. О времени возникновения мифа есть некоторые сведения - древние китайцы знали о нем, по меньшей мере, не позднее III в. до н.э.: в это время он уже нашёл отражение в китайской поэзии. Вот отрывок из стихотворения «Призывание души» Цюй Юаня, жившего в III в. до н.э.: Восточной стороне не доверяйся. Там великаны хищные живут И душами питаются людскими. Там десять солнц всплывают в небесах И расплавляют руды и каменья, Но люди там привычны ко всему. Таковы стихи III в. до н.э. о той стороне, где современное коренное население все ещё помнит легенду о десяти солнцах, нарушивших порядок поочерёдного появления на небе и вспыхнувших все разом. Итак, I тысячелетие до н.э., причём не позднее III в., а скорее всего, первая половина-середина I тысячелетия - такова историческая датировка этногенеза тунгусо-маньчжурских народов Приморья, которую можно сопоставить с данными генохронологии. Генетическое исследование тунгусо-маньчжурских народностей Приморья было проведено в 1970-1973 гг. под руководством проф. Ю. Г. Рычкова приморской исследовательницей В. Г. Ворониной. ЗАСЕЛЕНИЕ АМЕРИКИ. Перейдя рубеж нашей эры, мы оказались в раннем железном веке в пору его расцвета повсюду в Евразии, в том числе на Дальнем Востоке. Пожалуй, только крайний северо-восток нашей страны в это время все ещё пребывает в каменном веке в отношении материальной культуры. Весь образ жизни этих племён и даже тип их жилища, как указывал академик А. П. Окладников, оставались и на пороге новой эры близким к образу жизни древнейшего населения Сибири - охотников на мамонтов и на других крупных стадных животных ледниковой эпохи. Исчезнувших мамонтов, носорогов, лошадей заменили киты, моржи, тюлени - добыча, которая по-прежнему надолго обеспечивала пишей все население какого-либо посёлка на побережье пролива и моря Беринга. Поэтому без особых потерь информации пойдём ещё дальше в глубь времени - в верхний палеолит. Это отнюдь не пора безлюдья и запустения. Костры на стоянках верхнепалеолитических охотников светились повсюду на территории нашей страны. В том числе и в Сибири, которая не подвергалась покровному оледенению, где на месте современной равнинной и горной тайги и даже тундры простирались различные варианты степных ландшафтов. Именно в это время произошло подлинное - первое в истории человечества - открытие Америки древними сибирскими охотниками на мамонта. Следуя за стадами мамонтов, они пересекли тот условный меридиан, который сегодня разделяет два острова Диомида. В то время, о котором идет речь, вместо островов перед людьми, по-видимому, предстали две относительно небольшие, покрытые кустарниками сопки, каких и сегодня немало на Чукотке и Аляске. И к северу, и к югу от этих сопок простиралась суша, а реки Чукотки и Аляски продолжали течь в своих долинах по этой земле, как текут они и сегодня под водами Берингова моря. Первооткрыватели Америки шли по земле, которую после таяния ледников покрыло море. И о суше, ставшей дном Берингова и Чукотского морей, теперь догадываются лишь ученые, называя эту исчезнувшую землю Берингией. Но когда Сибирь и Америка ещё не были разделены морем, а Америка даже не была заселена человеком, предки тех, кого мы сейчас называем «коренным населением Америки» и «коренным населением Сибири» генетически отличались не более, чем сегодня различаются популяции в пределах одного народа. Радиоуглеродных дат для стоянок, найденных российскими и американскими археологами, много, и разброс в оценках времени велик. Но все же средняя дата (по данным на 80е годы XX века, когда было проведено описываемое исследование) для самых древних стоянок статистически вполне реальна: 25±3.4 тысяч лет назад. Эта дата и включена в генохронологию. Ею отмечен канун такого события, как разделение исторических путей популяций Америки и Сибири. НАРОДЫ СИБИРИ. Оставим теперь в стороне праамериканцев и обратимся к населению, оставшемуся в Сибири и начавшему формировать собственный генофонд, то есть к далеким прямым предкам современных сибирских народов. Эпоха верхнего палеолита в укладе жизни этих людей ещё продолжалась. Очень медленно, растянутые на тысячелетия, происходили незаметные изменения климата, ландшафта, животного мира в направлении к современным. Такие изменения едва ли могут быть отмечены в памяти сотен поколений. Воспоминания о них накапливаются лишь в виде мифов и воспринимаются как чудесные сказки. Лишь современная археология может установить их здравый смысл. Лишь современная генетика может подтвердить, что столь далекие предки действительно существовали и более того - их гены дошли и до сказителя, и до слушателей его сказок. Радиоуглеродные датировки существуют для многих сибирских стоянок различной древности. Здесь можно провести лишь условную границу древности, основываясь уже не на географических рубежах вроде Берингова пролива, а на рубежах различных археологических культур. Тогда для древнейших стоянок средняя радиоуглеродная дата (по данным на 80е годы XX века) составит 20 ± 2.5 тысяч лет назад. Люди, оставившие для последующей радиоуглеродной датировки угли костра, зажжённого в ледниковом периоде, передали и другие вести в будущее, и, прежде всего потомков, разведших в тех же или в новых местах и в новых поколениях новые костры. Добавим в заключение, что генетическое исследование современных коренных народов Сибири и Америки проведено соответственно российскими и американскими генетиками и антропологами за последние 20-30 лет XX века. По отношению к тем десяткам тысячелетий, на которые нам предстоит опуститься в глубь времен, эти два-три десятка лет, конечно, могут быть приняты за единую точку отсчёта времени. Как мы видим, в представленных фрагментах истории народов нет никакой системы. Единственное условие их включения в геохронологический анализ - наличие в прошлом какого-либо события, время которого документировано и которое могло иметь генетические последствия на всем дальнейшем протяжении времени, вплоть до современности, в которой мы пробуем отыскать его генетические следы. РАСЧЁТ ГЕНЕТИЧЕСКИХ ДАТ Для этого генохронология использует простую формулу: Ft= Fe(1-e-t/2Ne) =(4NeMe+1)-1(1-e-t/2Ne), где t - символ неизвестного нам числа прошедших со времени событий поколений, Fe =(4NeMe+1)-1, то есть определяется селективно-нейтральными демографическими данными о размере популяций Ne и миграций генов Мe (раздел 4 Приложения.). Формула описывает накопление с ходом времени межпопуляционных различий Ft. Эта величина различий между популяциями, достигнутая по прошествии t поколений от разделении прапопуляции, представляет собой ныне наблюдаемые нами генетические различия между дочерними популяциями Ft= FST. Таким образом, левая часть равенства Ft=FST оценивается нами как средняя FST=L-1∑FST(i) по данным о L множестве i-тых генов, каждый из которых может быть подвержен тому или иному типу отбора. Это означает, что генофонд данной исследуемой группы населения был изучен по широкому спектру (i) генетических маркёров. Для каждого i-ro маркёра были получены свои оценки - FST(i) - дифференциации популяций в пределах генофонда данной группы населения. Далее оценки FST(i), были усреднены по всей совокупности i-тых генов и для данного генофонда получена средняя оценка: FST. Правая же часть равенства содержит только селективно-нейтральные параметры - время t, размер популяций Ne, миграции генов Ме. На основании этого равенства, предполагающего селективную нейтральность FST, рассчитываются оценки генохронологии - время в поколениях t, прошедшее от исходного исторического события до времени изучения этого генофонда. Время в поколениях t было умножено на среднюю величину поколения (25 лет), что позволило перейти к более привычным для нас датам солнечного календаря - в годах. Если гипотеза верна - исторические датировки должны быть близки к генетическим датировкам. Если гипотеза неверна - между историческими и генетическими датами сходства не будет. СРАВНЕНИЕ ГЕНЕТИЧЕСКИХ И ИСТОРИЧЕСКИХ ДАТ Итоги генохронологии сведены в таблице 4.1. Сопоставим описанные выше исторические даты (левый столбец), полученные по данным истории и археологии, с датами генетическими (правый столбец), полученными по данным об изменчивости современного генофонда. ХОТОНЫ. Историческая дата: постановление о переходе хотонов от земледелия к скотоводству было принято в 1934 г. Генетический календарь показал, что для генофонда хотонов переломным стал год 1936. УЙГУРЫ и ДУНГАНЕ. По историческим данным дунгане и уйгуры переселились из Китая в 1870-1884 гг. Генетическая датировка: 1870-1886 гг. АЛЕУТЫ. Исторически зафиксированное заселение алеутами Командорских островов приходится на 1826-1829 гг. Генетическая датировка: 1827 г. 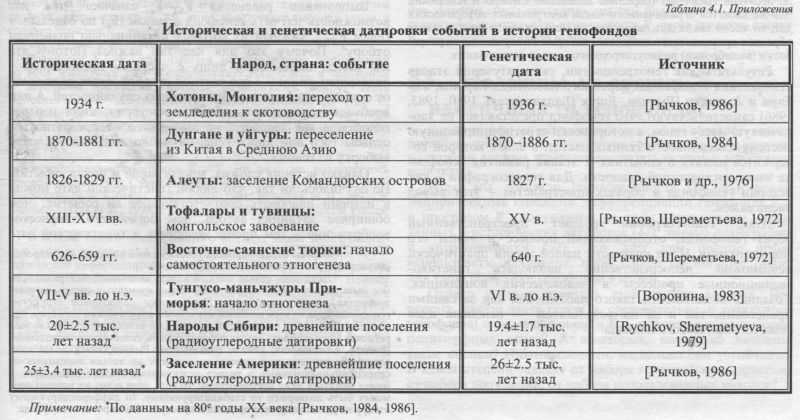
ТОФАЛАРЫ и ТУВИНЦЫ. Исторические летописи дают двухвековой разброс для даты «рассечения» золотыми приисками земли тофаларов (XIII—XVI вв.). Генетическая датировка легла в эти пределы: 1420 г. ВОСТОЧНО-САЯНСКИЕ ТЮРКИ. По историческим данным, начало самостоятельного этногенеза восточно-саянских тюрок, вероятнее всего, приходится на период 626-659 гг. Генетическая датировка: 640 г. ТУНГУСО-МАНЬЧЖУРЫ ПРИМОРЬЯ. Начало этногенеза тунгусо-маньчжуров Приморья по генетической датировке определяется VI в. до н.э. Не эти ли события, вызвавшие вспышку этногенеза, привлекли внимание Конфуция (551-479 гг. до н.э.), совершившего путешествие к «восточным иноземцам»? НАРОДЫ СИБИРИ. Средняя радиоуглеродная датировка древнейших поселений (20±2.5 тыс. лет назад) перекрывается с датировкой генохронологии (19.4±1.7 тыс. лет назад), полученной по оценкам дифференциации сибирского генофонда, включающего все современные коренные народы Сибири. ЗАСЕЛЕНИЕ АМЕРИКИ. И, наконец, что касается времени открытия и заселения Америки, то здесь генетические часы (26 тыс. лет назад) вновь показывают такую же точность, что и радиоуглеродные (25 тыс. лет назад). Отметим ещё одно важное обстоятельство. Радиоуглеродные часы указывают на время появления археологических стоянок, на время, когда на этих стоянках горели костры (ведь угли от древних костров и используются главным образом для такой датировки). Но что стало в дальнейшем с людьми, оставившими эти стоянки? Об этом, конечно же, радиоуглеродный метод не позволяет судить. Генетические же часы говорят и об этом. Показываемая ими дата означает, что гены тех людей, что разожгли эти костры на своих стоянках тысячи лет назад, дошли до нашей современности. Итак, все генетические датировки случайно выбранных исторических событий в происхождении самых разных народов оказались соответствующими датировкам историческим. Совпадение датировок открывает удивительную возможность анализа древней истории населения по данным о его современном генофонде, о генетической дифференциации ныне живущих популяций. Конечно, генохронологические датировки немногочисленны и в этом отношении оставляют сомнение, как всякое ограниченное наблюдение. Однако эта ограниченность иного рода, чем та, с которой обычно мы встречаемся при статистических наблюдениях. Те исторические события, которые были подвергнуты генохроно-логическому изучению, рассеяны почти по всему мыслимому диапазону времени человеческой истории - от десятков лет (хотоны) до десятков тысяч лет (коренное население Сибири и Америки). Что же касается ограниченного числа контрольных исторических дат, то число таких дат оказалось не меньше числа контрольных образцов органического вещества, которые послужили для разработки (калибровки) радиоуглеродного метода датирования. Результаты как генохронологии, так и изучения этапов становления генофондов коренного населения Европы, Сибири и Америки [Рычков, Ящук (Балановская), 1980, 1983, 1986] свидетельствуют, что генофонд представляет не хаотическую массу генов, а исторически стратифицированную систему изменчивости генных частот, в слоях которой содержится память о событиях и этапах развития генофонда многотысячелетней давности. Для геногеографии - как истории генофонда в образах пространства - этот вывод очень важен. Итак, геногеография создаёт пространственный образ генофонда, отображающий процесс и этапы его формирования. Из-за краткости нашей жизни практически невозможно непосредственно наблюдать генетико-эволюционные процессы в человеческих популяциях. Создание возможности такого наблюдения как за самими процессами, так и за их результатом — основная идея геногеографического подхода. §2. Соответствуют ли данные генетики прогнозу?Итак, мы выявили согласованность между историческими датами и датами генетическими, полученными по множеству генов, подверженных отбору. Что нам это дает? Прежде всего, согласованность дат, полученная для огромного диапазона человеческой истории - от современности до палеолита - говорит о точности генетической летописи, хранимой нашим генофондом. Свидетельствует о ясной и глубокой памяти генофонда. Во-вторых, она свидетельствует об объективности геногеографии, что является предметом обсуждения данного раздела. Но самое главное для предмета всей нашей книги и возможности изучения генофонда - такая согласованность свидетельствует о селективной нейтральности средних генетических различий между популяциями FST, полученных по множеству селективно-значимых генов. Это положение лежало в основе генетических датировок: FST≈Fe=1/(4NeMe+1). Напомним, что Fe - это величина селективно-нейтральной дифференциации генофонда. Это тот ожидаемый уровень генетических различий между популяциями, который определяется демографическими параметрами размера популяций и миграций Fe=1/(4NeMe+1)-1 при стационарном процессе3, то есть когда достигнуто равновесие между дрейфом и миграциями. Стационарные распределения являются базисными во многих аспектах изучения генетической структуры популяций. Например, согласно Р. Левонтину: «... популяционная генетика - это теория, рассматривающая равновесное состояние...», причём «... в практических приложениях мы пользуемся только равновесными положениями и стационарными распределениями» [Левонтин, 1978; стр.275]. При этом стационарность генетического процесса (например, при использовании равенства Fe=[4NeMe+1]-1) обычно принимается по умолчанию. Эмпирически наблюдаемые различия между популяциями FST оцениваются как средняя арифметическая по всему набору (L) i-тых генетических маркёров, то есть FST=[L-1∑FST(i)], где i=1, 2..., L. Близость оценок FST≈Fe связана с тем, что с ходом времени FST растёт не линейно, а по экспоненте. Большая часть генетического разнообразия (40-80% от Fe) возникает уже в самые первые поколения существования дочерних популяций, как это было показано Ю. Г. Рычковым для популяций Берингоморья и R. Ward для индейцев племени яномама (см. [Рычков, 1984]). Выполнение равенства FST≈Fe означает для нас возможность изучить генофонд в целом (Fe) по отдельным генам FST(i), каждый из которых потенциально подвержен отбору4. Почему это для нас так важно? Потому что мы всегда имеем дело лишь с частным проявлением - отдельным геном и с его частной судьбой, столь зависимой от особенностей среды и многих иных случайностей. А нам необходимо разглядеть общее - «исторический» портрет генофонда как такового, не зависящего от превратностей судьбы того или иного гена, случайно попавшего в нашу выборку из генома. Однако история сложна, многогранна и полна событий. Не случилось ли так, что любой генетической дате можно в истории подобрать соответствующее ей событие? Чем обширнее эрудиция автора, чем богаче его творческое воображение - тем проще подобрать к генетической дате её исторический эквивалент. Как ответить на такого рода сомнения? Любые ответы - что чем дальше вглубь, тем меньше исторических дат; что генетический смысл имеют лишь те исторические события, которые привели к переломному изменению популяционной структуры, и так далее - звучат как оправдание и оттого не убедительно. Тем более, что нас самих всегда отпугивали смелые интерпретации генетического рельефа в терминах истории - слишком велик соблазн отыскать в калейдоскопе исторических событий такое, которое может объяснить особенности географического распространения гена. Поэтому попробуем найти другие ряды доказательств равенства FST=Fe, пусть не столь эффектные, как генохронология, но зато более прямолинейные. Логика здесь проста. Надо сравнить не даты, а непосредственно эмпирические данные генетики (FST) и их прогноз -величину Fe, получив прогноз из качественно иных данных, не имеющих отношения к генетике. Такие «не генетические» и вообще «не биологические» данные нам могут дать две науки. Первая наука - демография. С помощью чисто демографических оценок размера популяций Ne и миграций Me, можно получить искомый прогноз Fe=1/(4NeMe+1). Вторая наука -лингвистика, или точнее антропонимика - наука об именах и фамилиях. Используя данные о фамилиях как об аналогах генов, можно получить независимые, не биологические и потому не зависящие от отбора (селективно-нейтральные) оценки дифференциации генофонда Fe. Обе оценки - по демографии и по фамилиям - дают прогноз величины Fe по «не генетическим» данным. Получив этот прогноз, мы можем сравнить его с эмпирической величиной FST, полученной по данным генетики. Выражение FST≈Fe=1/(4NeMe+1) означает, что средние оценки FST, полученные по случайной репрезентативной выборке классических или ДНК маркёров, соответствуют селективно-нейтральному уровню дифференциации генофонда Fe. Этот вывод явился эмпирически обобщением, полученным в результате анализа многочисленных данных самыми разными исследователями. При этом выявлено, что случайная выборка из генома размером 20-30 локусов (или 50 аллелей) уже дает устойчивую оценку FST [Cavalli-Sforza, Piazza, 1993; Bowcock et al., 1987; Айала, Кайгер, 1988; Fuerst et al., 1977; Chacraborty et al., 1978; Nei, 1975]. Например, в работах L. L. Cavalli-Sforza и его коллег [Bowcock et al., 1991a; Bowcock et al., 1991b] считается достаточным для корректной оценки FST≈Fe лишь увеличение выборки генов, а среднее значение FST рассматривается как селективно-нейтральное без каких-либо дополнительных обоснований. Равенство FST≈Fe принимается по умолчанию практически во всех известных нам исследованиях. Например, в обширной и детальной дискуссии, которая развернулась вокруг теста на селективную нейтральность Левонтина-Кракауэра (где Fe определяется через FST) и в которой участвовали крупные авторитеты в области популяционной генетики (R. С. Lewontin, J. Krakauer, М. Nei. Т. Maruyama, A. Robertson, W. J. Ewens, M. W. Feldman, L. B. Jorde, L. L. Cavalli-Sforza и др.) основополагающее равенство FST≈Fe ни разу не подвергалось сомнению [Nei, 1965; Cavalli-Sforza, 1966; Lewontin, Krakauer, 1973; Левонтин, 1978; Кимура, 1985; Jorde, 1981; Bowcock et al., 1991a,b; Sanchez-Mazas et al., 1994; Poloni et al., 1995; Cavalli-Sforza, Piazza, 1993]. В основе такого общепринятого подхода лежат многочисленные данные, полученные при сопоставлении генетической и не генетической информации о дифференциации генофонда. Например, в основополагающей монографии L. L. Cavalli-Sforza. W. F. Bodmer [1971] приводятся результаты изучения широкого спектра генов в различных регионах ойкумены: в подразделённой популяции Италии, в коренном населении Африки и у аборигенов Новой Гвинеи. Авторы приходят к заключению о выполнении равенства FST≈Fe: «во всех этих случаях F оценки, полученные по данным о генных частотах, и ожидаемые F оценки, полученные на основе матрицы брачных миграций, находятся в хорошем согласии» [Cavalli-Sforza, Bodmer. 1971, стр. 454]. Целенаправленно и пристально вопрос о равенстве FST≈Fe рассматривался в многочисленных работах Ю. Г. Рычкова и его школы. Одна из наших совместных работ по этнической генетике [Рычков, Балановская, 1990 а] специально была посвящена обобщению всей накопленной информации о сравнении эмпирических оценок FST и селективно-нейтрального показателя Fe, получаемого по «не генетическим» данным. Результаты, приведенные в таблице 4.2. убедительно свидетельствуют о близости эмпирических оценок FST и их прогноза Fe, полученного по «не генетическим» данным - как по демографии, так и по фамилиям. При этом равенство FST≈Fe оказывается необычайно устойчивым и выполняется при разных, столь варьирующих условиях (табл. 4.2.): а) при использовании для оценки FST значительно различающихся панелей генов; б) при анализе популяций разного происхождения (от Прибалтики до Дальнего Востока); в) при анализе популяций разных иерархических уровней и древности - от современных народов до Сибиро-Американской общности, уходящей корнями в верхний палеолит. Остановим внимание лишь на одном удивительном моменте: равенство FST≈Fe соблюдается даже в совокупности циркумполярных популяций Заполярья, включающей целый ряд народов Арктики - от лопарей на западе до алеутов и эскимосов на востоке [Рычков, Шереметьева, 1976; Rychkov, Sheremetyeva, 1979]. Эти популяции в течение длительного времени преемственного развития (по-видимому, ещё с донеолитической эпохи, 10-5 тыс. до н.э. [Рычков, Шереметьева, 1976; Rychkov, Sheremetyeva, 1979]) испытывали мощное давление экстремальных условий среды, чрезвычайно суровой и практически однородной во всем огромном ареале Заполярья. Естественно было бы ожидать, что именно в этих популяциях мощное давление однородной среды отклонит оценки разнообразия FST от селективно-нейтрального уровня Fe и приведёт к нарушению равенства FST≈Fe Однако этого не произошло! Показанное сохранение равенства FST≈Fe даже в подразделённой популяции, находящейся в столь экстремальных условиях и занимающей обширнейшую территорию (все Заполярье Восточного полушария площадью 3750 км2), указывает, что в менее экстремальных ситуациях мы тем более можем определять селективно-нейтральную изменчивость Fe через среднюю оценку FST по репрезентативной выборке генов. 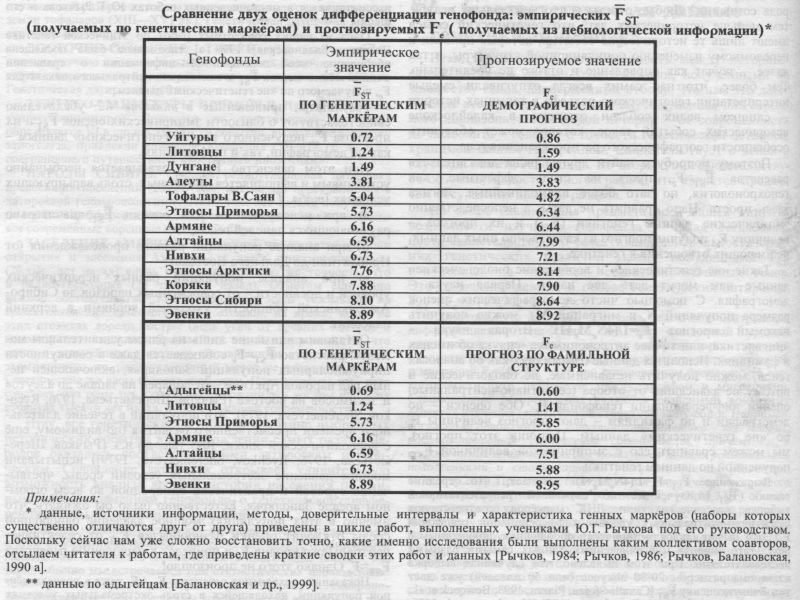
§3. Надёжны ли оценки изменчивости генофонда?В этом разделе мы рассмотрим вопрос об устойчивости оценок изменчивости генофонда и попробуем сформулировать те правила, выполнение которых позволяет рассчитать надёжные устойчивые величины. Итак, мы доказали согласованность (FST≈Fe) между эмпирическими оценками дифференциации генофонда FST и прогнозом Fe, полученном по трем независимым рядам данных - исторических датировок (§1), данных демографии и антропонимики(§2). Это дает нам полное право использовать обобщение, проведённое по выборке генов, для характеристики генофонда в целом. Однако следует ответить ещё на два вопроса. Во-первых, какую часть генов (а, следовательно, и тотального генофонда) описывают оценки FST, получаемые по выборке полиморфных генов? А во-вторых, насколько надёжны такие оценки? Иными словами, насколько они устойчивы? В какой степени зависят от набора генетических маркёров, от набора популяций, от набора использованных методов? Специально проведённые исследования [Балановская, Рычков, 19906,в; Рычков, Балановская, 1990а] позволили сформулировать два вывода. ВЫВОД 1 Средние оценки различий между популяциями FST≈Fe, полученные по классическим и ДНК маркёрам, адекватно описывают межпопуляционное разнообразие структурных генов генома. При этом они не зависят от того, получены ли они по классическим (иммуно-биохимическим) маркёрам или же по ДНК маркёрам (молекулярно-генетическим). Согласно [Bowcock et al., 1987; Рычков, Балановская, 19906; Bowcock et al., 1991a,b; Cavalli-Sforza, Piazza, 1993] с помощью таких оценок FST мы описываем средний уровень изменчивости аутосомных генов - наиболее значительной части генома, включая не только уникальные последовательности ДНК, но и мини- и микросателлитные последовательности [Лимборская и др., 1998; Беляева и др., 1998]). Отметим, что факт независимости FST от типа маркёров -классические или ДНК - до сих пор порой вызывает недоумение. Ход рассуждения таков. Как же так? Классические маркёры определяются по их белковым продуктам, и потому должны быть намного сильнее подвержены отбору, чем ДНК маркёры - случайно взятые фрагменты ДНК, в большинстве своем от отбора укрытые. Из этого делается, казалось бы, логический вывод, что изменчивость (FST) классических маркёров, ограничиваемая отбором, должна быть намного меньше, чем изменчивость ДНК маркёров. Ошибка «логики» здесь в том, что она помнит только о стабилизирующем типе отбора, сужающим изменчивость (FST), забывая о дифференцирующем отборе, который может увеличить изменчивость гена (FST) по сравнению с селективно нейтральной. Если же выборка генов случайна по отношению к отбору - то отбор не в силах сместить среднюю оценку FST. Данные, приведённые в таблице 4.3. для мирового генофонда (и подробнее рассмотренные в разделе 8.1.) демонстрируют очень хорошее согласие между классическими и ДНК маркёрами в оценках различий между популяциями. Причём для восточноевропейского генофонда анализ проведён максимально строго. Во-первых, и ДНК, и классические маркёры анализируются в Восточной Европе по полностью идентичному набору этносов. А это очень важный момент. Представим, что в одну выборку народов Восточной Европы мы бы включили только восточных славян, а в другую - и восточных славян, и лопарей, и калмыков, и народы Кавказа. Понятно, что реальных размах генетических различий во второй выборке должен быть намного шире, чем в первой (чисто «славянской»). И тогда разница между уровнями изменчивости двух типов маркёров могла бы быть вызвана не тем, что изучены разные маркёры, а тем, что изучены разные наборы народов. Во-вторых, в выборку ДНК маркёров включены, главным образом, высокополиморфные мини- и микросателлитные маркёры. Это привело к тому, что уровень гетерозиготности Hs оказался в два раза (!) выше, чем по классическим маркёрам. И, несмотря на это, оценки межпопуляционных различий FST оказались одинаковы {раздел 7. 1.). Такое соответствие оценок FST по классическим и ДНК маркёрам - при идентичном наборе популяций и резко различном размахе полиморфизма (гетерозиготности) - указывает на то, что мы получаем не частную оценку изменчивости конкретной выборки генов, а общую оценку изменчивости генофонда. ВЫВОД 2 Средние оценки различий между популяциями генофонда (FST≈Fe) обладают высокой устойчивостью. В таблице 4.3. приведены четыре ряда оценок разнообразия мирового генофонда, полученных принципиально различными методами [Levontin, 1972; Latter, 1980; Рычков, Балановская, 1990 б; Bowcock et al., 1987]. Все ряды оценок резко различаются по всем исходным параметрам: по числу изученных народов мира и их составу; по числу генных маркёров и их составу; и даже сами меры межпопуляционного разнообразия - различны. Общими для этих работ были лишь охват всего мирового разнообразия в целом и проведение исследования на едином (этническом) уровне популяционной системы. При этом оценки межпопуляционного разнообразия F оказались чрезвычайно устойчивыми и индифферентными к методическим расхождениям авторов. Характер маркёров - белковые продукты генов [Levontin, 1972; Latter, 1980; Рычков, Балановская, 1990 а] или же ДНК-полиморфизм [Bowcock, 1987; Bowcock et al., 1991a; Bowcock, Cavalli-Sforza, 1991] - также не сказался на значениях FST и HS (табл. 4.3.). Полученный результат является важнейшим свидетельством устойчивости средних оценок разнообразия генофонда. 
НАСКОЛЬКО УСТОЙЧИВЫ ОЦЕНКИ ДИФФЕРЕНЦИАЦИИ ГЕНОФОНДА? Основываясь на приведённых данных, кратко ответить на этот вопрос можно так. Средние оценки дифференциации генофонда FST, полученные по репрезентативному набору полиморфных маркёров (и по классическим, и по ДНК маркёрам) характеризуют основную часть генофонда. Эта характеристика отражает селективно-нейтральный процесс формирования генофонда. При корректной организации материала (выборка генов, выборка популяций, выборка индивидов), эта характеристика является чрезвычайно устойчивой, что можно считать отражением реального уровня изменчивости генофонда. Однако, сужая масштаб исследования - переходя от генофонда всей ойкумены, всего человечества ко всё более дробным уровням популяционной структуры, - мы переходим к исторически менее устойчивым генофондам и должны быть готовы к менее устойчивым оценкам генного разнообразия. Здесь от исследователя требуются особая методическая точность и чуткость, чтобы компенсировать сужение масштаба исследований. Это особенно важно, когда мы переходим к анализу лишь одного этноса. Не претендуя на полноту, попробуем обобщить методические требования к изучению генного разнообразия в подразделённых популяциях меньшего размера, чем ойкумена. ТРЕБОВАНИЕ ПЕРВОЕ: ЕДИНЫЙ УРОВЕНЬ ПОПУЛЯЦИОННОИ ИЕРАРХИИ Это требование означает, что нельзя включать в один анализ, например, данные о народах в целом (среднеэтниче-ские) и данные о локальных популяциях внутри этноса. Иерархичность организации генофондов не требует специальных доказательств. Определим популяцию человека как исторически сложившуюся, занимающую определённый ареал и устойчивую в поколениях совокупность людей, генетический вклад которых в каждое последующее поколение своей популяции больше, чем вклад мигрантов (пришельцев из иных популяций). Такой генетический вклад (соотношение генов из «своей» популяции и из «иных» популяций среди всех генов, переданных в следующее поколение) рассчитывается на основе брачных миграций и оценивается через гаметный вклад. Кроме генофонда человечества, то есть видовой популяции Н. sapiens (гаметный вклад которой по определению 100%, поскольку нет межвидовых скрещиваний), этому определению наиболее бесспорно отвечают этносы - племёна, народности, народы, нации. Этнографы и антропологи знают: если в каком-либо этносе начинают устойчиво преобладать браки с пришельцами из иных народов, значит, дни этого этноса могут быть сочтены. Этнический уровень организации популяционной иерархии существует как данная нам реальность: сами этносы, уровень их эндогамии, их история, динамика их ареалов определены достаточно рельефно, благодаря этническому самосознанию [Долуханов, 2000]. Именно поэтому выбор как единиц исследования именно этносов (а не административных или территориальных групп населения) позволяет сравнивать примерно на едином уровне иерархии даже столь разные генофонды, как, например население Европы и коренные народы Америки [Рычков, Ящук (Балановская) 1980, 1982, 1983, 1984,1986; Rychkov, Yashchuk (Balanovskaya), 1985]. Для проведения исследования на иных уровнях популяционной иерархии (выше или ниже этноса) требуется реконструкция популяций с помощью разных наук - демографии, этнографии, лингвистики, антропологии. Даже при выделении наиболее очевидного уровня элементарных популяций необходимо учесть гаметный вклад. Показано, что даже в разных частях ареала одного и того же этноса - русского народа - элементарным популяциям соответствуют разные группы населения: отдельные деревни (Архангельская, Вологодская области), сельсоветы (Костромская область), группы сельсоветов (Кировская область), районы (Белгородская область) и даже группы районов (Краснодарский край) [Пасеков, Ревазов, 1975; Ревазов и др., 1979, 1988; Сладкова и др., 1990; Брусинцева и др., 1993; Сорокина. 2005]. При изучении подразделенного генофонда необходимо стремиться к тому, чтобы все изучаемые субпопуляции принадлежали к одному уровню популяционной иерархии - будь то элементарные популяции или этносы. Нарушение этого требования может исказить оценки межпопуляционного разнообразия. Как бы ни было трудно фиксировать уровни популяционной иерархии, ещё больше осложнений возникнет, если их не выявить, если включить в анализ популяции безотносительно к их рангу в популяционной системе. Поясним на примере. Даже в крупных исторически сложившихся популяционных системах (таких, как коренное население Америки, Сибири, Европы) третья часть всей межпопуляционной изменчивости FST (рис. 4.1.) приходится на разнообразие популяций в пределах этноса [Рычков, Ящук (Балановская), 1980. 1982, 1983, 1984, 1986; Rychkov, Yashchuk (Balanovskaya), 1985]. 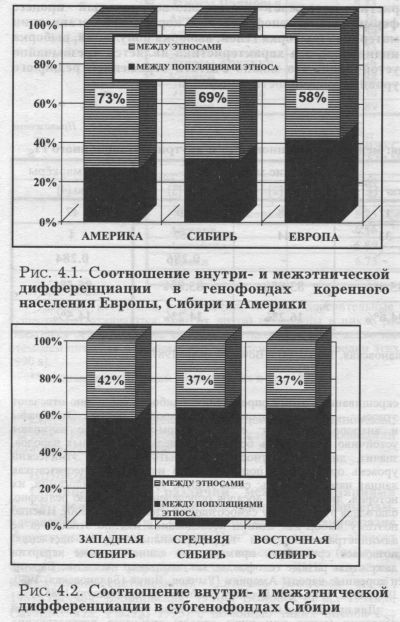
Теперь спустимся на один популяционный уровень - к субрегионам (рис. 4.2.) и рассмотрим три субрегиона Сибири - Западную, Среднюю и Восточную Сибирь. Мы видим, что в них на разнообразие популяций в пределах этноса приходится уже много больше - две трети от всей межпопуляционной изменчивости FST субрегиона (рис. 4.2.). Предположим, что исследование Средней Сибири будет проведено на уровне этносов, а Восточной Сибири - на уровне локальных популяций. Из рис. 4.2. ясно, что будет нельзя сравнивать полученные оценки изменчивости! Мы видим, что дифференциацию Средней Сибири в этом случае мы занизили в три раза (откинув 63% всей изменчивости, которая приходится на различия локальных популяций внутри этноса) по сравнению с Восточной Сибирью. Например, исследователь №1 решил сравнить два субрегиона - Среднюю и Восточную Сибирь. При этом он совершил ошибку смешения разных уровней иерархии, смешения разных «матрёшек»: в Средней Сибири - оценил различия между этносами, а в Восточной Сибири - между локальными популяциями. Иными словами, он оценил разнообразие генофонда Средней Сибири (обозначим как MS) по различиям между его народами -долганами, нганасанами, кетами, энцами и другими. Он включил в анализ средние частоты для каждого народа (этнический уровень). В этом случае он получил бы в результате величину FST(MS - №1)=2.595. А в Восточной Сибири (ES) исследователь №1 включил в анализ у бурят - все 17 изученных популяций, а не одну «средне-бурятскую» частоту, для эвенов - включил все 15 изученных популяций вместо «средне-эвенской», для якутов - все 36 популяций, для юкагиров - все 4 популяции. И так далее. То есть в Восточной Сибири он провёл анализ на другом уровне - уровне различий локальных популяций в регионе, а не этносов. Поэтому он получил бы в результате величину FST(ES - №1)=6.37. Сравнив изменчивость двух субрегионов Сибири, исследователь №1 пришёл бы к выводу, что разнообразие генофонда Средней Сибири (2.59) намного меньше, чем Восточной Сибири (6.37): FST(MS - №1) << FST(ES - №1) Предположим, что исследователь №2 также решил сравнить эти два субрегиона. Он совершил ту же ошибку смешения уровней иерархии, смешения разных «матрёшек», только как бы наоборот: в Средней Сибири - оценил различия между локальными популяциями, а в Восточной Сибири - между этносами. То есть, рассматривая генофонд Средней Сибири, он включил в анализ все 8 популяций долган вместо «средне-долганской» (как делал исследователь №1), все 4 популяции нганасанан вместо «средне-нганасанской», все 5 популяций кетов, все 4 популяции энцев и т. д. Иными словами, в Средней Сибири он провёл анализ на уровне локальных популяций, а не этносов. Тогда он получил бы в результате величину FST(MS - №2)=7.00. А в Восточной Сибири исследователь №2 поступил включил в анализ у нивхов Сахалина одну «средне-нивхскую» частоту, для эвенов - «средне-эвенскую», для якутов - «средне-якутскую», для юкагиров -«средне-юкагирскую». И так далее. То есть в Восточной Сибири он провёл анализ на уровне не локальных популяций, а этносов. Тогда он получил бы в результате величину FST(ES - №2)=2.36. Сравнив изменчивость, исследователь №2 пришёл к выводу, что разнообразие генофонда Средней Сибири (7.00) намного больше, чем Восточной Сибири (2.36): FST(MS - №2) >> FST(ES - №2)! Итак, оба исследователя изучали одни и те же регионы, использовали одну и ту же базу данных, исходные данные у них были совершенно одинаковы. Но выводы их оказались диаметрально и статистически достоверно (!) противоположны. И при этом оба вывода неверны. На самом деле изменчивость этих двух генофондов практически одинакова, причём на любом из уровней иерархии! На уровне различий между популяциями субрегионов в Средней Сибири FST(MS)=7.00. а в Восточной Сибири FST(ES)=6.37. На уровне различий между этносами субрегионов в Средней Сибири FST(MS-ET)=2.59, а в Восточной Сибири FST(ES-ЕТ)=2.36. А единственная причина, по которой получены совершенно неверные выводы лишь в том, как они организовали одни и те же данные, на каком уровне популяционной иерархии проводили анализ - причина в смешении разных уровней иерархии, сравнении разных «матрёшек». Поясним ешё одним примером - реальным. Авторитетный исследователь6 изучил один из коренных народов Сибири, и получил величину различий между популяциями этого народа FST(данного этноса) ≈ 9. Много это или мало? Чтобы ответить на этот вопрос, он сравнил с величиной изменчивости популяций Сибири FST(Сибири) ≈ ll. И сделал вывод, что различия популяций этого этноса несколько меньше, чем это принято в Сибири, но, в обшем-то, такие же, как для других сибирских народов. Но вся беда в том, что та величина, с которой он сравнивал свой единственный этнос, FST(Сибири) ≈ ll, охватывает популяции не одного этноса, а все популяции всех этносов Сибири! Иными словами, в эту величину (FST(Сибири) ≈ ll) уже вошли не только различия между популяциями усредненного сибирского этноса, но и различия между всеми народами всей Сибири. На рисунке рис. 4.1. мы видим, что средняя гетерогенность сибирского этноса (различия между популяциями одного «усредненного» народа Сибири) составляет треть от FST(Сибири) ≈ ll, то есть примерно FST(среднего этноса) ≈ 3. Это означает, что народ, изученный этим исследователем, обладает уникально огромными различиями между популяциями - в три раза большими, чем для среднего этноса Сибири! Что требует специальных исследований и объяснений. Налицо совершенно неверный вывод - гетерогенность этого сибирского этноса (на изучение которого было потрачено столько сил!) вовсе не меньше среднего по Сибири (как заключил автор), а в три раза больше «нормы»! И столь ошибочный вывод был сделан лишь потому, что автор забыл об уровнях популяционной иерархии. Ошибка FST, заданная неправильной организацией материала, столь велика, что сопоставление генофондов теряет всякий смысл: FST зависит уже не от свойств генофонда, а лишь от соотношения этносов и локальных популяций в выборке. Если наиболее изученные регионы (или гены) исследованы на уровне локальных популяций, а менее изученные - на уровне этносов, то оценки FST(i) - только за счет выборки популяций - для наиболее изученных объектов будут на треть-две трети больше, чем для слабо изученных. В этом случае сравнение регионов (или генов) будет отражать только ошибку в организации данных, а не особенности генофонда. Сравнительное изучение дифференциации региональных генофондов или разных генов в этом случае проблематично: оценки абсолютно ненадёжны, независимо от того, каковы статистические оценки их достоверности. Поэтому, сравнивая дифференциацию разных генов FST(i) при анализе одного и того же генофонда, мы должны включать в анализ популяции одного и того же иерархического уровня. Когда мы переходим к сравнению генофондов разных регионов, то здесь добавляется ещё одно требование: иерархические уровни самих регионов, самих тотальных популяций должны быть идентичны. Например, корректно сравнивать по величине дифференциации (FST) генофонды таких регионов, как Европа, Сибирь и Америка - они находятся на одном иерархическом уровне. Можно сравнивать субрегионы - генофонды Средней Сибири и Северной Европы -они также находятся на одном иерархическом уровне (субрегионов). Но нельзя сравнивать субрегион с регионом - например. Среднюю Сибирь с Европой в целом, считая их равноценными партнерами сравнения, забывая об их иерархии. Это наглядно видно из сравнения рис. 4.1. и 4.2. - легко рассчитать, что дифференциация FST Сибири в два раза больше, чем дифференциация её субрегионов. Значит, сравнивая Среднюю Сибирь и Европу, мы сравниваем «половинку» с «целым», приравниваем их. Что же сказать о таких сравнениях, как дифференциация отдельного этноса и региона в целом? В этом случае малая часть (треть или даже четверть) как бы приравнивается к целому! Выводы таких сравнений - а их немало в популяционных публикациях - содержат столь крупную методическую ошибку, что она сводит на нет все усилия авторов. Иными словами, сравнение популяций разных иерархических уровней содержит в себе очень серьёзную ошибку и приводит к заведомо неверным результатам, если при интерпретации мы забываем о разном «ранге» сравниваемых популяций. ТРЕБОВАНИЕ ВТОРОЕ: ПОЛИМОРФИЗМ ГЕНЕТИЧЕСКИХ МАРКЕРОВ Это требование налагает запрет на включение в анализ редких аллелей. Надёжность эмпирических оценок FST(i) зависит от уровня полиморфизма изучаемых генов. Эта важно учитывать при оценке селективных сил, поскольку при низком уровне полиморфизма (то есть если частота аллеля близка к 0 или 1) динамика частот генов «будет независима не только от далекого прошлого, но, возможно, и от недавней истории процесса отбора» [Левонтин, 1978]. Традиционно предлагаются два критерия полиморфизма: 1% (то есть 0.01< q <0.99) и 5% (0.05< q <0.95). Иногда - например, при исследовании гетерозиготности Н - вводятся и более жесткие рамки: Н>0.02, что соответствует 0.14< q<0.86. Необходимость введения критерия полиморфизма при анализе FST-статистик убедительно показана при компьютерном моделировании FST(i) при разных значениях генных частот (i) (рис. 4.3., приводится по [Bowcock et al., 1991]). Из графика видно, что при средней частоте гена в тотальной популяции q < 0.05 оценки межпопуляционной изменчивости FST(i) варьируют случайным образом в чрезвычайно широком диапазоне. Они выходят на плато при q = 0.05 и практически неизменны при q > 0.10. Вспомним, что для достоверного определения FST(i) и FST(i) при малых концентрациях гена необходимы ещё и огромные популяционные выборки: чем меньше частота аллеля, тем больше должна быть выборка для того, чтобы эта частота достоверно отличалась от нуля. Из этого становится ясно, как важно следовать критерию полиморфизма, и сколь высока возможность ошибки при включении генов с низким уровнем полиморфизма (редких аллелей). 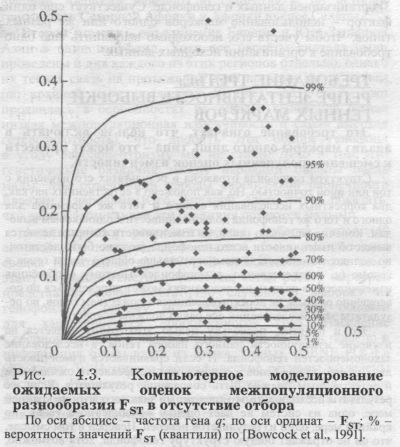
Насколько велика может быть эта ошибка, видно, например, из двух опубликованных оценок дифференциации коренного населения Австралии по гену FY*B: 1) FST(i)=0 при q=0.00 [Балановская, Рычков, 1990]; 2) FST(i)=8.5 при q=0.01 [Cavalli-Sforza, Piazza, 1993]. Величина FST(i)=8.5 очень велика - она составляет половину изменчивости человечества (FST(i)=14.5, табл. 4.3). Насколько можно судить по приведенным данным, во второй работе в выборку попали метисы коренных австралийцев с европейцами (у которых частота этого гена существенна) - такой случайности достаточно, чтобы на порядок изменить не реальную дифференциацию коренных австралийских популяций, а получаемую оценку дифференциации низко полиморфного гена. Итак, включение в выборку низко полиморфных генов приводит к неустойчивости и непредсказуемости оценок дифференциации генофонда FST, а также к отклонению от изменчивости по селективно-нейтральному типу (то есть FST ≈ Fe). Именно поэтому для получения устойчивых оценок дифференциации генофонда FST желательно использовать выборки генов, соответствующию строгому критерию полиморфизма: 0.05< q <0.95. Это важно ещё и потому, что для слабо полиморфных генов велика не только случайная ошибка. На её фоне проявляется и систематическая ошибка: включение слабо полиморфных генов вызывает занижение средней оценки дифференциации FST. Поясним этот момент. На рис. 4.4. представлены оценки для шести основных регионов мира из сводки [Cavalli-Sforza et al., 1994]. Мы рассчитали их в двух вариантах: 1) по всем маркёрам (0 Итак, рассмотрение разных оценок изменчивости генофондов основных регионов мира выявило, что устойчивость оценок FST связана с тремя факторами: 1) долей низко полиморфных генов; 2) средним уровнем их полиморфизма; 3) организацией данных о генофонде. Существует ещё один фактор - использование маркёров одного или же разных типов. Чтобы учесть его, необходимо выполнить ещё одно требование к организации исходных данных. ТРЕБОВАНИЕ ТРЕТЬЕ: РЕПРЕЗЕНТАТИВНОСТЬ ВЫБОРКИ ГЕННЫХ МАРКЁРОВ Это требование означает, что нельзя включать в анализ маркёры одного лишь типа - это может привести к смещению полученных оценок изменчивости. Структура генофонда отражена в результатах его изучения с той или иной точностью. Но, как положено в естественных науках, два корректных исследования одного и того же параметра для одного и того же генофонда должны привести к одинаковым выводам. Конечно же, число генов, по изменчивости которых делается вывод об изменчивости всего генофонда, должно быть достаточно велико, но оно всегда намного меньше общего числа генов в геноме (и, соответственно, в генофонде). Поэтому обобщённая изменчивость в разных исследованиях может определяться по совершенно разным, не перекрывающимся множествам генов, но результаты этих исследований все равно должны совпасть. Это и понятно, поскольку целью этих работ является не изучение изменчивости данного набора генов, а исследование закономерностей генофонда. И если сравнивается изменчивость двух разных типов генов в одном и том же ареале, то ожидаемым и естественным должно быть совпадение результатов. Если же результаты несходны, то это тревожный знак того, что, по крайней мере, одна из систем не дает оценку «обобщённого гена», по крайней мере, один из результатов неверен. Неверен в том смысле, что он не отражает общую структуру генофонда, хотя возможно, вполне удовлетворительно описывает изменчивость данного набора генов. Это снова подводит нас к вопросу о том, сколько и каких конкретных генов надо включить в анализ, чтобы их совокупная изменчивость соответствовала изменчивости некоего «обобщённого гена», отражала изменчивость не конкретного набора генов, а генофонда в целом. Но прежде ещё раз подчеркнём, что характеристику генофонда можно получить по разным панелям генетических маркёров, и все эти оценки могут быть правильными и равноценными и, что наиболее существенно, они должны совпадать друг с другом. Формулируя требования к репрезентативности выборки генов - по отношению ко всему геному - важно учитывать обе её стороны, количественную и качественную. 1) КОЛИЧЕСТВЕННЫЙ АСПЕКТ ФОРМИРОВАНИЯ ВЫБОРКИ - ЧИСЛО ГЕННЫХ МАРКЁРОВ В ДАННОЙ ПАНЕЛИ. Разные авторы пришли к эмпирическому обобщению, что, как правило, выборка примерно из 20-30 локусов (или =50 аллелей) является достаточной для надёжной оценки FST. При дальнейшем увеличении выборки средняя оценка FST практически не меняется [Bowcock et al., 1987; Айала, Кайгер. 1988; Cavalli-Sforza, Piazza, 1993]. Меньшая выборка генов тоже может давать корректные результаты, но необходима проверка её корректности. Меньшая выборка требует обязательного контроля её репрезентативности с помощью прямых оценок Fe (§2) или иных доказательств её адекватности и соблюдения равенства FST=Fe. 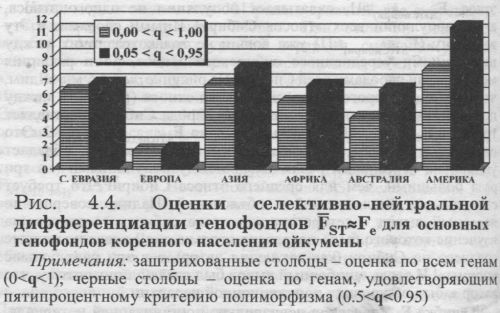
2) КАЧЕСТВЕННЫЙ СОСТАВ ВЫБОРКИ ГЕНОВ ИЗ ГЕНОМА - ЕЁ СЛУЧАЙНОСТЬ ПО ОТНОШЕНИЮ К ДЕЙСТВИЮ ЕСТЕСТВЕННОГО ОТБОРА. Если в выборке будут преобладать гены, подверженные стабилизирующему отбору - средняя оценка FST будет занижена. Если будут преобладать гены, на которые действует дифференцирующий отбор средняя оценка FST будет завышена по сравнению с истинной, селективно-нейтральной дифференциацией генофонда. При этом важно знать, что некоторые типы маркеров - например генетико-биохимические маркёры (в отличие от иммунологических) [Балановская, 1998] или, тем более, гены наследственной патологии - всегда занижают среднюю оценку FST, поскольку для них преобладающим является стабилизирующий отбор. В разных разделах мы приводили целый ряд примеров высокой устойчивости FST и выполнения равенства FST=Fe при использовании очень разных наборов классических маркёров. Адекватные выборки генов различаются столь сильно и по их размеру, и по их составу, что невозможно определить иное «правило» для их создания, чем случайность по отношению к эффектам отбора (что обычно обеспечивается включением в выборку самых разных типов классических маркёров). Более того, новые аутосомные маркёры не меняют картины: накапливающаяся _информация по ДНК маркёрам указывает, что их средние FST близки к FST=Fe, определяемым по «классическим» маркёрам (см., например, табл. 4.3. и раздел 8.1.). При включении в выборку новых маркёров со своеобразной функцией в организме, с особым характером микроэволюции, требуется специальная проверка выполнения для них равенства FST≈Fe. Такая проверка была проведена, например, для генов, связанных с иммунным ответом, для системы HLA (human leukocytal antigen) [Евсеева, 2001]. Ряд тесно сцепленных локусов этой генетической системы обладает выраженным полиморфизмом: панель аллельных вариантов каждого локуса необычайно обширна, поскольку обеспечивает генетический контроль иммунного ответа. Одни и те же народы Крайнего Севера, представляющие три разных лингвистических семьи, были изучены как по панели аллелей HLA (проверялось наличие 160 аллелей 6 локусов HLA-I и HLA-II классов), так и по панели классических маркёров (15 аллелей 5 локусов иммуно-биохимического полиморфизма). Каждый аллель HLA природой предназначен для ответа на запросы особой внешней среды. Поэтому можно было ожидать, что оценки дифференциации FST системы HLA - причём в экстремальных условиях Крайнего Севера! - будут чрезвычайно смещены (FST≠Fe) от селективно-нейтральных. Однако проверка показала, что - вопреки ожиданиям - средняя оценка дифференциации по системе аллелей HLA соответствует селективно-нейтральной дифференциации! Видимо, разнообразие факторов среды, на которые система HLA обеспечивает иммунный ответ, столь велико, что все разнонаправленные векторы компенсирует и взаимно «гасят» друг друга при расчёте средних оценок FST. В силу исключительного высокого полиморфизма система HLA дает устойчивую и объективную картину различий между генофондами. §4. Сравнение основных регионов мираПри рассмотрении русского генофонда мы учитывали характеристики генофонда ойкумены и генофондов коренного населения всех крупных историко-географических регионов мира. Поэтому кратко сравним их (табл. 4.4. и 4.5.). Эти характеристики, полученные в цикле работ Ю. Г. Рычкова и Е. В. Балановской, являются наиболее полным обобщением данных о генофонде всех регионов мира по классическим маркёрам, и поэтому служат важнейшим дополнением к обобщающей монографии Кавалли-Сфорца с соавторами [Cavalli-Sforza et а., 1994]. ОРГАНИЗАЦИЯ ДАННЫХ Эти характеристики получены по репрезентативной выборке классических маркёров (49 аллелей 20 локусов) и с учетом иерархической структуры популяций - в каждом из регионов в качестве популяций выступают этносы. Всего по частотам генов в ≈50000 популяций были получены характеристики 5135 этносов, в среднем по каждому локусу изучено 257 народов мира. Для всех видов анализа и во всех генофондах выполнены единые требования: репрезентативность и единообразие выборки генов для всех генофондов; достаточный уровень полиморфизма; единый уровень анализа популяционной системы (этнический); единый исторический масштаб и целостность генофондов (историко-географический регион). Организованный таким образом генофонд в наибольшей степени отвечает как требованиям, предъявляемым к объектам популяционно-генетического анализа, так и основным положениям и моделям теоретической популяционной генетики (в частности, предположению о стационарности генетического процесса). Во-первых, генофонд ойкумены является сложно организованной популяционной системой самого высокого уровня и потому обладает наибольшей устойчивостью средних генных частот во времени. Во-вторых, этносы являются не только наиболее точно фиксированными и универсальными популяционными единицами, но и сами обладают высокой устойчивостью средних генных частот. В-третьих, историко-географические регионы - в отличие от чисто географических, политических, расовых, культурологических или конфессиональных подразделений - наиболее полно и комплексно отражают вехи в пространственно-временной эволюции генофонда Homo sapiens. Они не позволяют оставить за рамками анализа те группы населения, которые по демографическим параметрам или темпам социально-экономического развития находятся на периферии современности (поскольку вклад популяции в генофонд человечества не определяется её численностью и динамичностью на данном временном срезе). Пояснения требуют лишь два момента. Историко-географические регионы охватывают все коренное народонаселение ойкумены за исключением населения Северной Африки: им пришлось пожертвовать, поскольку его включение в регион, объединяющий народы Европы и Юго-Западной Азии, могло показаться слишком смелым решением - также, как и выделение его в отдельный регион Средиземноморья в целом (Северная Африка, Южная Европа, Ближний Восток); а объединение с населением «черной Африки» неверно по существу - по истории генофондов Северной Африки и Африки южнее Сахары. Второй момент - объединение Европы и Юго-Западной Азии в один генофонд. Хотя все расчёты сначала были проведены и для каждого из этих регионов отдельно, однако их тесная связь на протяжении всей истории Homo sapiens (по крайней мере, последних 40 тысяч лет) столь ярко проявилась в особенностях их общего генофонда (самая низкая межпопуляционная изменчивость и самая высокая гетерозиготность), что дробить этот общий генофонд в угоду европоцентризму мы не стали. Дело в том, что генетические различия между народами Европы очень малы. Мы объединили народы европейской оконечности Евразии с народами Юго-Западной Азии, чтобы показать, что даже в этом случае эти различия остаются малыми. Анализируется репрезентативная выборка генов (49 аллелей 20 локусов). Для анализа была использована сводка [Mourant et al., 1976]. В ней подведен итог тому этапу изучения генетики человека, когда реестр маркёров представлял собой достаточно случайную выборку из генофонда и потому мог служить надёжной основой для характеристики «обобщённого» гена. Население Северной Евразии в сводке практически не представлено и информация о нем взята из Банка данных «GENE POOL» (см. главу 3). Историко-географические регионы выделены согласно [Mourant et al., 1976; Брук, 1981]. На основе популяционных частот 49 аллелей мы рассчитали частоты для каждого из народов мира. 
Основные характеристики генофондов мира сведены в таблице 4.5. Дадим только краткое описание некоторых параметров (подробно см. [Балановская, 1998]). ГЕНЕТИЧЕСКИЕ РАССТОЯНИЯ «РЕГИОН - МИР» Степень сходства по частотам генов между каждым из регионов с одной стороны и ойкуменой с другой стороны определена через генетические расстояния dРЕГ-МИР (рис. 4.5., табл. 4.5.). Диапазон различий оказался чрезвычайно велик: он достигает трёхкратного уровня, колеблясь от 0.108 в Северной Евразии до 0.360 в Австралии. Коренное население Австралии, Африки и Меланезии в пространстве генных частот значительно удалено от мировых средних (dРЕГ-МИР > 0.24), то есть эти регионы генетически своеобразны, они являются «генетическими окраинами» ойкумены. Ближе находятся Евразийские регионы (В.Азия, Индостан и Европа), и наиболее близок к мировому «генетическому центру» (dРЕГ-МИР=0.l08) генофонд Северной Евразии. 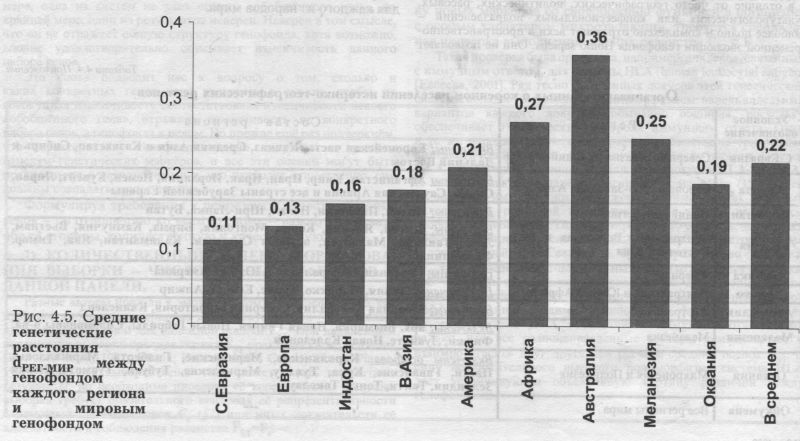
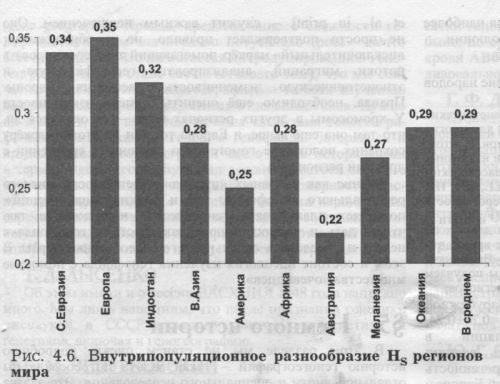
ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ Для каждого генофонда рассчитаны (табл. 4.5.) три основные характеристики_генного разнообразия [Nei. 1975] в среднем по всем генам: HT-общее разнообразие региона, HS- средняя гетерозиготность этносов (рис. 4.6.); GST~FST -дифференциация этносов в регионе. Мы видим, что уровень гетерозиготности HS в коренном населении трех регионов достоверно выше, чем в среднем по всем генофондам: он выше в Европе, Северной Евразии, Индостане (Hs>0.32). Достоверно снижена гетерозиготность коренного населения Австралии и Америки (HS<0.25). Уровень межэтнических различий FST концентрирует в себе итоги микроэволюции генофонда, связанные с эффектами генных миграций и дрейфа генов: размах разлbчий по FST между регионами во много раз выше, чем по HS и HT (табл. 4.5'.). Достоверно выше среднерегионального FST в Америке и Меланезии, достоверно ниже - в населении Европы и В.Азии. Население Северной Евразии характеризуется высоким уровнем всех показателей генетического разнообразия: по уровню общего разнообразия HT этот генофонд занимает первое место в мире; по уровню гетерозиготности HS - второе; по уровню FST - третье место в мире. 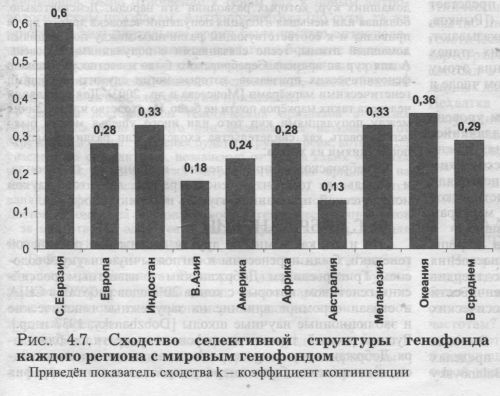
СЕЛЕКТИВНАЯ СТРУКТУРА Направление и интенсивность отбора оценивались через отклонения FST(i) от селективно-нейтрального уровня FST. Рассмотрен новый параметр генофонда — его селективная структура, в которой выделяются три класса генов. Класс «LOWER DIFF» включает гены, дифференциация которых достоверно ниже селективно-нейтральной: FST(i)<FST; для этих генов выдвигается гипотеза стабилизирующего отбора. Класс «NEUTRAL» включает условно-нейтральные гены с межпопуляционной изменчивостью FST(i), достоверно близкой к селективно-нейтральной: FST(i))≈FST. Класс «SUPER DIFF» включает гены, дифференциация которых достоверно выше селективно-нейтральной: FST(i)>FST; для этих генов выдвигается гипотеза дифференцирующего отбора. На основе трех статистических критериев — χ2, t, F - была определена достоверность отличий FST(i)) от FST для каждого гена в каждом из генофондов ойкумены и получены оценки селективной структуры генофондов. Показано, что в разных генофондах одни и те же гены подвержены разным типам отбора. Наименее устойчив класс «NEUTRAL»: ни один из генов не оказался селективно-нейтральным во всех регионах мира. Генный состав класса «LOWER DIFF» в генофондах мира более стабилен, чем класса «SUPER DIFF». Степень сходства по селективной структуре в целом (коэффициент контингенции k) региональных генофондов с мировым позволяет распределить генофонды по степени близости к «общевидовому» ответу на давление среды (табл. 4.5., рис. 4.7.). В целом по структуре отбора регионы имеют незначительное сходство с общемировым (k=0.29). В восточно-азиатском (k=0.18) и австралийском (k=0.13) генофондах сложились свои независимые типы селективной структуры генофондов. Генофонды коренного населения Африки и Европы, Америки и Индостана, Меланезии и Океании, столь различные и географически, и антропологически, - демонстрируют один и тот же невысокий уровень сходства с глобальной селективной структурой (k≈0.3). И лишь один регион - Северная Евразия - занимает особое положение: уровень его сходства с глобальной селективной структурой (k=0.60) в два раза выше среднерегионального! При этом высокий уровень сходства отмечается не для какого-нибудь одного, а для всех классов селективной структуры: для LOWER DIFF коэффициент связи k=0.6, для NEUTRAL k=0.5, для SUPER DIFF k=0.7. ИТОГИ. СЕВЕРНАЯ ЕВРАЗИЯ Сведем воедино показатели, полученные с помощью трех основных инструментов - частот генов, их разнообразия и селективной структуры (табл. 4.5, рис. 4.5.-4.7.). Комплексная оценка позволила получить принципиально новые результаты. Одним из них является особое положение генофонда Северной Евразии - по всем независимым показателям он наиболее полно воспроизводит мировой генофонд. Из всех региональных генофондов мира лишь генофонд Северной Евразии занимает центральное положение в пространстве как генных частот (наименьшее расстояние до мирового генофонда dРЕГ-МИР= 0-108), так и селективной структуры (наибольший показатель сходства с мировой селективной структурой kРЕГ-МИР), и сохраняет наибольшее генное разнообразие (HT=0.362). Поэтому именно этот регион является наиболее информативным для изучения общих проблем эволюции. ИТОГИ. ЕВРОПА Вопрос о том, каково генетическое разнообразие народов Западной Европы, обычно волнует всех. В главе 5 (табл. 5.3.1.) мы привели оценки генетических различий между локальными популяциями (FПОП-ЭТН) для многих народов Западной Европы и для многих народов Сибири. И в том, и в другом регионе рассмотрены наиболее генетически изученные народы. По одним и тем же наиболее изученным классическим маркёрам (ABO, РТС, MNS, RH, FY, FUT2, PI, KEL, LE, HP, GC, TF) мы рассчитали среднюю генетическую гетерогенность для европейского этноса и для сибирского этноса (FПОП-ЭТН - различия популяций внутри одного народа). Сводка данных по Сибири была составлена нами и Н. А. Таусик в то же время, что и сводка по Зарубежной Европе [Mourant et al., 1976]. «Выровняв» таким образом и число народов, и число аллелей, мы получаем наиболее сопоставимые оценки для генофондов этих регионов. Различия популяций в пределах этноса Зарубежной Европы оказались чрезвычайно малы: FПОП-ЭТН=0.76±0.24. Средние генетические различия популяций в пределах коренного сибирского этноса очень велики: FПОП-ЭТН=4.04±0.98. Это означает, что гетерогенность сибирского этноса в пять раз выше, чем в Западной Европе. Обратим внимание, что эта величина почти в три раза меньше, чем приведённая в таблице 4.5. FЭТН-РЕГ=2.41 для «синтезированного» нами много более гетерогенного региона, объединившего Западную Европу с Юго-Западной Азией. Рассчитаем теперь генетические различия не внутри этноса, а между народами Западной Европы - может быть, они окажутся велики? Нет, генетические различия между теми же народами Западной Европы столь же малы и составили всего лишь FЭТН-РЕГ=0.87. Теперь мы можем рассчитать, каковы же генетические различия между всеми популяциями всех народов Западной Европы. Этот общий размах межпопуляционных различий для Европы составил: FПОП-РЕГ = FПОП-ЭТН + FЭТН-РЕГ = 0.76 + 0.87 = 1.71. Таким образом, оказывается, что все генетические различия всего населения Западной Европы (FПОП-РЕГ=1.71) в два раза меньше, чем гетерогенность (FПОП-ЭТН=4.04) лишь одного «среднего» коренного народа Сибири! Таким, образом, генетические различия всех элементарных популяций всей Зарубежной Европы - от португальцев до лопарей и от исландцев до греков - достигают лишь чрезвычайно низкого уровня. В генетическом масштабе населения не только ойкумены, но и всех иных регионов мира, европейский мир предстает генетически гомогенным. Наши исследования ([Рычков, Ящук (Балановская), 1983; Балановская, 1998]) показывают, что эта гомогенность возникает не на последних этапах развития европейской цивилизации, а свойственна этому региону практически на всем протяжении его, в том числе и не писаной, истории. Но в данном контексте важно установить сам уровень генетических различий популяций Европы, являющийся итогом особенностей её исторического развития. Важно и то, что эти результаты получены по классическим маркёрам четверть века назад. Эти результаты позволяли прогнозировать и столь же низкое генетическое разнообразие народов Западной Европы по маркёрам митохондриальной ДНК. Широко развернувшиеся в последние годы исследования мтДНК народов Европы показали удивляющую многих гомогенность населения Западной Европы (см. главы 6, 8, 9), полностью подтвердив возможность прогнозирования параметров изменчивости большинства ДНК маркёров по данным о классических маркёрах. Большинства - но, к счастью, не всех. Высокая изменчивость гаплогрупп Y хромосомы - как в пределах Европы, так и в пределах русского народа [Balanovsky et al., in print] - служит важным исключением. Оно не просто подтверждает правило, но и обнаруживает «исключительный» маркёр, помогающий дифференцировать потоки миграций, анализировать географическую и этноисторическую изменчивость населения Европы. Правда, необходимо ещё оценить степень изменчивости Y хромосомы в других регионах мира - не окажется ли, что там она ещё выше, и Европа тогда и по этому маркёру сохранит положение гомогенного региона (в сравнении с другими регионами). Знание как основных «правил» изменчивости каждого регионального генофонда, так и редких «исключений», позволяет планировать генетическое исследование так, чтобы дать и полноценную характеристику генофонда в целом, и проследить отдельные его своеобразные черты. В этом и состоит идеология изучения генофонда с помощью множества «очевидцев». §5. Немного историиВ этом разделе мы не станем писать подробную историю геногеографии - такая задача потребовала бы отдельной книги и специального исследования. Это задача скорее для историков науки, а наше видение, конечно же, субъективно. Но все же и наши сведения имеет смысл привести - наравне с иными свидетельствами они помогут восстановить истинную картину. К тому же мы лишь наметим основные вехи развития нашей науки - в том свете, как они видятся авторам. Причём опишем их далеко не всегда в хронологическом порядке, а следуя логике развития науки. Для удобства чтения назовём эти вехи по именам ученых, внесших решающий вклад на поворотных этапах геногеографии, а для современного этапа будем считать такими вехами разработку того или иного геногеографического метода. А.С. СЕРЕБРОВСКИЙ Возникновение и термина «геногеография», и самой идеи чтения истории популяций в книге пространственного распространения генов связаны с именем Александра Сергеевича Серебровского. В этой книге мы уже не раз упоминали его имя как основателя геногеографии. Поэтому сейчас приведём только один пример элегантности его работ: едва ли не первым геногеографическим исследованием было изучение истории народов Дагестана по географической изменчивости морфологических признаков у домашних кур, которых разводили эти народы. Действительно, большая или меньшая изоляция популяций человека друг от друга приводит и к соответствующим различиям между популяциями домашней птицы, тесно связанными с популяциями человека. А для кур во времена Серебровского было известно уже немало фенотипических признаков, которые могли служить хорошими генетическими маркёрами [Моисеева и др., 2003]. Для популяций человека таких маркёров почти не было. Но сходство или различия между популяциями кур того или иного ущелья можно было использовать как свидетельства сходства или различий между популяциями их хозяев. Серебровскому принадлежит и понятие генофонда, и мысль о том, что геногеография является наукой исторической, призванной изучать историю генофонда. Ф. Г. ДОБРЖАНСКИЙ Эти идеи, как и многие другие достижения российской генетики, были перенесены в англоязычную науку Феодосием Григорьевичем Добржанским - известным россий¬ским генетиком, который с конца 20х годов работал в США и оказал огромное влияние на зарубежные генетические и эволюционные научные школы [Dobzhansky, 1937 и др.]. Русскоязычный термин «генофонд» превратился благодаря Добржанскому в «gene pool», аналогично на американском континенте была интродуцирована и геногеография (gene geography). Правда, представление о сложности генофонда при этом значительно потускнело (pool - нечто бесструктурное). Что же касается интродуцированной геногеографии, то она стала соответствовать тому, что мы сейчас называем частной геногеографией - то есть изучению географического распространения отдельных генов. Нужно ли говорить, что при этом исчезла сама тема геногеографического изучения не генов, а генофонда. Затерялось и представление о геногеографии как о науке исторической - термин «gene geography» стал ассоциироваться скорее с изучением медицинских следствий из закономерностей распространения гена, чем с изучением истории народов. Мы перечислили эти особенности зарубежного восприятия геногеографии, чтобы ярче подчеркнуть те черты этой науки, которые особенно важны для отечественной традиции. Т. Д. ЛЫСЕНКО Об этом имени и о сессии ВАСХНИЛ 1948 года написано много. Мы лишь напомним, что после признания генетики лженаукой в СССР начались гонения на генетику и генетиков, включая и геногеографию. Е. М. ЧЕПУРКОВСКИЙ Однако геногеографический или очень близкий к нему по духу подход сохранялся в среде антропологии, которая, хотя и ценой больших жертв, избежала участи генетики (см. об этом в разделе 2.1). Антропологию того времени связывали с геногеографией два моста. Первым был географический подход, который постепенно становился краеугольным камнем этнической антропологии. Этот подход, восходящий к классическим работам Ефима Михайловича Чепурковского, можно в терминах популяционной генетики описать так: для популяций, сходных по значениям ряда признаков и занимающих соседние географические ареалы, можно предполагать их родство по происхождению; а популяции, сходные по значениям ряда признаков, но географически далекие друг от друга, нельзя считать родственными по происхождению, если для этого нет иных убедительных свидетельств. Этот географический метод важен для антро-пологии тем, что он отметает множество фантастических гипотез, основанных на сходстве нескольких признаков в популяциях, которые географически и исторически удалены друг от друга. В. В. БУНАК Вторым мостом, связывающим антропологию с гено-географией, было отношение к генам как к новым антропологическим признакам. Антропогенетика (по сути, та же геногеография) длительное время была частью антропологии, и ведущие антропологии возлагали большие надежды на использование этих новых признаков («кровяных групп») - ведь они лишены многих недостатков, свойственных «обычным» антропологическим признакам. Главным достоинством генетических признаков в глазах антропологов было то, что они проявляются независимо от окружающей среды: у ребенка будет та группа крови, которую он унаследовал от родителей, независимо от того в каких условиях ребенок рос. А для многих антропологических признаков на генетическую, унаследованную составляющую, накладывается влияние условий жизни индивидуума и всей популяции. Достоинством была и «дискретность» генетических признаков - за них отвечал один ген, а не совокупность генов, как для антропологических признаков. Поэтому таким разочарованием проникнута работа ведущего антрополога Виктора Валерьяновича Бунака [Бунак, 1969]. Ее основной вывод - в «полиареальности», то есть мозаичном, а не закономерном распределении частот генов по ареалу. Это ведь сейчас мы знаем, что распределение генов в действительности подчиняется строгим закономерностям - но чтобы их выявить, нам необходимо проанализировать десятки генов. А в распоряжении антропологов того времени было, по сути, только две генетические системы - группы крови АВО и резус, поэтому и был получен результат «полиареального» распределения генов. Г. Ф. ДЕБЕЦ Столь глубокая внутренняя связь антропологии и гено-географии и была причиной того, что после снятия в нашей стране запрета на генетику геногеография возродилась из антропологических недр. Это второе рождение геногеографии связано с именем Юрия Григорьевича Рычкова, а связующим звеном, пронесшим мечту о настоящей генетической антропологии через десятилетия запрета на генетику, был его учитель, ведущий антрополог советской школы Георгий Францевич Дебец. По словам Юрия Григорьевича, мечтой и целью Дебеца было создание со временем подробного атласа распространения генетических признаков в населении нашей страны. И когда эта работа стала возможной (как по политическим, так и по техническим условиям), она была начата под руководством Ю. Г. Рычкова. Ю. Г. РЫЧКОВ С именем Юрия Григорьевича связаны широкомасштабные исследования генофонда народов СССР, которые проводились в рамках генетико-антропологической экспедиции не только в Сибири, но и во многих других регионах СССР. Но главное - именно благодаря Ю. Г. Рычкову началось возрождение геногеографии как науки. Для реализации мечты Г. Ф. Дебеца о геногеографическом атласе первым необходимым шагом было сведение воедино всех опубликованных и архивных данных о частотах генетических систем в популяциях СССР. Эта огромная работа была начата для населения Сибири Н. А. Таусик, завершала эту работу Е. В. Ящук (Балановская). На этом этапе первоочередным стал вопрос, который ставил ещё Г. Ф. Дебец - вопрос об организации данных для картографирования на популяционном или же. на этническом уровне. Для большинства антропологов естественно является анализировать (и картографировать) популяционные данные как они есть - то есть использовать имеющиеся значения признака для каждой изученной популяции и наносить на карту эти значения в точки, занимаемые популяциями. Альтернативный подход - усреднить значения во всех изученных популяциях данного народа и наносить на карту именно усреднённое, этническое значение признака, помещая его в исторический или географический центр этнического ареала. Для антропологических признаков обычно пользуются популяционными данными, но для генетических признаков был весомый аргумент в пользу картографирования этнических данных. Этот аргумент состоял в ограниченном объёме, в нехватке генетических данных для сплошного популяционного картографирования. Нехватка данных усугублялась тем, что большинство популяций были изучены по немногим генам, и наборы этих генов для каждой популяции различались (программа признаков, изучаемых антропологами, стандартизована в значительно большей мере). Поэтому наиболее объективную генофонда картину можно было создать именно на этническом уровне. АНАЛИЗ GST В ПОПУЛЯЦИОННЫХ СИСТЕМАХ. ЭКВИДИСТАНТНОСТЬ Но прежде, чем использовать вместо частот в локальных популяциях среднеэтнические частоты генов, необходимо было понять, не потеряется ли основная часть изменчивости при таком переходе от популяционных к этническим частотам? В более общем виде - какие закономерности связывают эти характеристики? Прекрасно разработанный в популяционной генетике аппарат анализа межпопуляционной изменчивости в иерархических системах популяций был применён к генетическим данным о народонаселении Сибири, а затем Западной Европы и Америки. Причём рассматривались не двууровневые системы («локальные популяции» и «этносы»), а многоуровневые системы (добавляя над уровнем «этносы» уровни «лингвистические группы» и «лингвистические семьи» при лингвистической классификации или иные объединения этносов при этноконфессиональной, антропологической и этнографической классификациях народов) [Рычков, Ящук (Балановская), 1980, 1983, 1986]. Результатом этих работ стало открытие принципа эквидистантности (равноудалённости) разных уровней популяционной системы: изменчивость популяций в пределах этноса была равной изменчивости этносов в пределах лингвистической группы, и межпопуляционная изменчивость средних частот аллелей в лингвистических группах, относящихся к одной лингвистической семье, равнялась все той же величине. Эквидистантность оказалась характерна не только для всех изученных популяционных систем человека (коренное население Сибири, Америки и Европы) [Рычков, Ящук (Балановская), 1984], но и для других видов организмов [Алтухов, 2003]. Была предложена (и доказана её справедливость) популяционно-генетическая модель скачкообразного роста популяционной системы, состоящего в распаде единой прапопуляции на дочерние, которые, в свою очередь, со временем распадаются на популяции следующего, низшего уровня. И при росте популяционной системы на один уровень, генетическое разнообразие популяций увеличивается на одну и ту же величину («квант»), что и приводит к феномену эквидистантности всех уровней системы [Рычков, Ящук (Балановская), 1985]. Феномен эквидистантности позволил дать и общий ответ на вопрос - можно ли строить карты по среднеэтническим частотам генов? Стало ясно, что в зависимости от масштаба, от ранга популяционной системы мы теряем при этом лишь около трети или четверти информации, но приобретаем устойчивость оценок. Этим открывалась возможность создания картографического атласа генофонда СССР. Разработанная тогда методология анализа межпопуляционной изменчивости широко используется в геногеографии и сейчас. Уровень межпопуляционной изменчивости является универсальным и весьма информативным показателем при характеристике любого этнического или регионального генофонда. Для генофондов крупных регионов методология применяется уже в полную силу - с анализом изменчивости на разных уровнях популяционной системы и с обязательным использованием феномена эквидистантности. Примером может служить изучение генофонда Кавказа [Балановская и др., 1999; см. также раздел 2.2. этой книги]. ПЕРВЫЕ КАРТОГРАФИЧЕСКИЕ ТЕХНОЛОГИИ Несмотря на успехи, достигнутые при чисто статистическом анализе генофондов, целью отечественной геногеографической школы оставалось создание крупных картографических атласов. При наличии исходных данных вопрос упирался в метод построения карт, который должен был быть, во-первых, объективным (воспроизводимым в руках разных исследователей), а во-вторых, обеспечивать «непрерывное» картографирование (расчёт ожидаемого значения частоты гена для каждой точки карты, даже вдалеке от непосредственно изученных популяций). Выбор метода интерполяции - способа расчёта ожидаемого значения из данных по опорным точкам - стал главной задачей. Поскольку интерполяционное картографирование необычайно трудоёмко и требует значительных вычислений, этот метод с самого начала задумывался как компьютерный. Однако на каждом этапе карты строились также и вручную - это позволяло параллельно развивать теоретические подходы к картографированию. В течение нескольких лет были созданы три разных компьютерных метода - триангуляционный, «чернильных пятен» и средневзвешенной интерполяции - и созданы соответственно три разных картографических технологии. Опишем их предельно кратко и образно. Триангуляционный метод разрабатывался совместно с сотрудниками кафедры биофизики МГУ А. Т. Терехиным и Е. В. Будиловой. Он основывается на соединении всех изученных популяций линиями, формирующими густую нерегулярную сеть треугольников, и вдоль этих линий вычисляются градиенты частоты гена. В результате из множества треугольников рождается визуальная карта распространения гена. К сожалению, у этого метода серьёзное ограничение - значение признака рассчитывалось для областей, и эти области менялись от карты к карте. Нельзя было рассчитать точные значения частоты гена для каждой точки карты. Исследователь видит результирующую карту, но не может получить числовую матрицу, лежащую в её основе. А значит, и не может дальше делать с картой любые статистические преобразования, не может превращать одну карту в другую или по совокупности исходных карт создать синтетическую карту. Поэтому следующим методом в геногеографии стала технология, созданная совместно с А. В. Рычковым. Ее образно можно назвать «технология чернильных пятен». Каждая изученная популяция представлялась в виде небольшого «пятна» с заданной интенсивностью цвета, соответствующей частоте гена. Начинался итерационный процесс, на каждом шаге которого каждое пятно, видимое на экране компьютера, расширялось на один пиксель. На каком-то этапе соседние пятна (происходящие из соседних изученных популяций) начинали сливаться, и тогда зона «контакта популяций» окрашивалась в промежуточный цвет или же в серию переходов промежуточных цветов, если популяции различались друг от друга более чем на один интервал шкалы. Полным завершением этого процесса было бы полное слияние всех пятен друг с другом, когда вся карта представлялась равномерным серым пятном - все генетические различия полностью нивелировались! Поэтому главным вопросом при использовании этой технологии был выбор - на каком шаге итерации следует остановиться, чтобы популяции успели «провзаимодействовать», но не стали бы полностью идентичными. Этот вопрос решался по анализу дисперсии на каждой итерации: дисперсия, изначально очень большая, постепенно снижалась и выходила на плато. Именно этот момент и был сигналом к остановке итерационной процедуры и рассмотрению полученной карты. Как можно видеть, такой метод по сравнению с триангуляционным обладает тем преимуществом, что позволяет рассчитать значения для каждой точки карты. Этот метод был использован для первого геногеографического атласа, созданного для населения Монголии [Балановская и др., 1990]. Однако дальнейшее развитие геногеографии оказалось связанным с иным, третьим методом, который также позволял получить числовые значения для каждой точки карты, но обладал и рядом дополнительных преимуществ. А главное - субъективные факторы (как, например, остановка на той или иной итерации) были сведены к минимуму. ТЕХНОЛОГИЯ НА ОСНОВЕ СРЕЛНЕВЗЕЩЕННОЙ ИНТЕРПОЛЯЦИИ Ведущая роль в создании этой технологии принадлежит картографической группе кафедры картографии МГУ под руководством С. М. Кошеля. Наша совместная работа является замечательным примером долгого и плодотворного сотрудничества профессиональных картографов и геногеографов. Еще до встречи с генетиками группа С. М. Кошеля широко использовала картографический пакет MAG и разработала оригинальную программу Metacopy для визуализации карт. Привнесение генетической методологии значительно изменило ситуацию. Программный пакет превратился в GG MAG (GG обозначает геногеографию - Gene Geography) и приобрёл множество функций, специально нацеленных на анализ генетических данных: начиная от использования данных в нерегулярно расположенных опорных точках (поскольку генетические данные именно таковы) до внедрения в пакет широких возможностей математических и статистических операций с картами. Пакет GGMAG прошёл длительную эволюцию, и в настоящий момент используется его последняя, четвёртая версия с некоторыми оригинальными добавлениями. Подробное описание самой технологии приводилось в разделе 3 Приложения, поэтому здесь укажем лишь её основные черты. Во-первых, при интерполяции для каждой регулярно расположенной точки карты (узел «сетки» карты) рассчитывается с помощью ортогональных полиномов средневзвешенное значение, получаемого из значений в нерегулярно расположенных опорных точках (популяциях). Значения во множестве опорных точках усредняются, причём каждая опорная точка выступает с весом, обратно пропорциональным расстоянию от нее до данного узла сетки. Тем самым прогнозируемое значение приближается к значениям в ближайших опорных точках, а удалённые опорные точки сказываются на прогнозе незначительно. Заметим, что тот же принцип независимо использовался и в нескольких зарубежных программах, специально созданных для картографирования генетических данных. Правда степень весовой функции от расстояния (между узлом сетки и опорной точкой) в них строго фиксирована (обычно это квадрат), тогда как в пакете GGMAG исследователь может задавать любую степень (часто оптимальные результаты получаются при использовании шестой или даже десятой степени). Другое важнейшее отличие GGMAG - использование для интерполяции ортогональных полиномов. Это позволяет адекватно прогнозировать изменение значений признака в пространстве даже при отсутствии данных, прямо указывающих на это изменение: интерполяционный алгоритм улавливает наличие тренда по окружающим точкам и продолжает его на смежные территории. Аналогично выбору степени весовой функции, исследователь может задать и различную степень полинома (при нулевой степени расчёт ведется без учета полинома, только как средневзвешенная интерполяция). Исследователь также может сам определить тот радиус, в пределах которого учитываются опорные точки (исходные популяции). Это позволяет моделировать разный круг взаимодействующих популяций. Сочетания всех трёх параметров (радиус для опорных точек, степень весовой функции расстояния до опорной точки, степени полинома) создают широчайший спектр возможностей для построения наиболее корректной компьютерной модели при данном сочетании популяций и значений признаков. КАРТЫ. ПОСТРОЕННЫЕ ВРУЧНУЮ Хотя магистральной линией было создание компьютерной геногеографии, на разных этапах множество карт было построено и вручную. С компьютерными картами их роднило всё - и непрерывность картографирования (значения для каждого узла равномерной сетки карты), и использование математического расчёта, а не научной интуиции при выборе картографируемого значения, при этом сам алгоритм вычислений мог быть самым разным. «Ручные» карты, сыгравшие наибольшую роль в истории геногеографии - это карты антропологических признаков в русских популяциях. Эта работа была специально задумана как проверка картографического метода - предполагалось, что если эти новые методы чего-то стоят, то они должны выявить новое даже в наиболее полных и прекрасно разработанных данных. А наиболее полными были данные по антропологии русского народа, собранные Русской антропологической экспедицией. Результаты этого геногеографического анализа антропологических данных описаны в разделе 2.3, поэтому скажем только, что успех этой работы превзошел ожидания. О следах летописных славянских племён, обнаруженных, благодаря геногеографическому методу в современном русском населении, был сделан доклад на авторитетной конференции, на которой присутствовало большинство отечественных антропологов и археологов [Рычков, Балановская, 1988]. То полное признание и одобрение, которое получили эти результаты, следует целиком отнести к тому стилю картографирования и той методологии анализа, которая впоследствии оформилась в компьютерную геногеографию. С этого времени можно начинать отсчёт широкого применения отечественной картографической технологии в изучении популяций человека. БАНКИ ДАННЫХ Наряду с картографической технологией и статистическими методами важное место в геногеографии занимают банки данных. Дело в том, что объём данных, которые используются даже не в слишком масштабном геногеографическом исследовании, огромен, а сами исходные данные обычно рассеяны во множестве статей. Представьте себе анализ даже такого сравнительно небольшого, хотя и гетерогенного региона, как Кавказ. Пусть Вам необходимо проанализировать данные только по классическим маркёрам. Однако подобные данные получали и публиковали самые разные авторы - от медицинских исследований групп крови АВО и резус, до работ по многим системам, выполненными в московских и кавказских генетических лабораториях. Некоторые данные по Кавказу публиковались и зарубежными авторами. Библиография данных о Кавказе насчитывает многие десятки наименований, и далеко не все эти статьи легко доступны, а многие из них даже неизвестны большинству исследователей. Итак, тому, кто решит провести геногеографическое изучение Кавказа, предстоит провести сперва большую работу по сбору и систематизации данных о частотах генов, опубликованных за несколько десятилетий. Если же представить, сколько разных генофондов было изучено геногеографами, и вообразить, какой объём предварительной работы требуется, к примеру, для анализа в масштабе СССР, то задача сбора и систематизации опубликованных генетических данных предстанет во всем своем величии и неприступности. Кардинальное решение этой задачи может быть только одно - собрать ВСЕ когда-либо опубликованные данные по частотам генов в ЕДИНОЕ хранилище. И при анализе любого генофонда лишь обращаться в хранилище и брать готовые систематизированные данные по частотам генов в популяциях интересующего региона. Именно таким хранилищем и стал банк данных «GENEPOOL», разработанный под руководством одного из авторов этой книги. В банк данных вводилась практически вся доступная информация по частотам классических маркёров в популяциях Северной Евразии, а отчасти и по другим регионам мира. Впоследствии банк был пополнен и данными по аутосомным ДНК маркёрам. Структурная организация банка и запрограммированные функции делают его не только хранилищем данных, но и инструментом их проверки, систематизации и анализа. Аналогичный банк данных создавался и зарубежной геногеографической школой Кавалли-Сфорца. Он содержит информацию по остальным регионам мира (кроме Северной Евразии, представленной весьма скудно), хотя запрограммированные возможности этого банка не столь обширны. Отечественной геногеографической школой создавался и ряд других банков данных: «ПАЛЕОЛИТ РОССИИ», база данных о русских фамилиях, банк данных по митохондриальной ДНК в населении мира и банк данных «РУССКИЙ ГЕНОФОНД» кратко описаны в разделе 5 Приложения. ЦЕЛЬ - СИНТЕТИЧЕСКИЕ КАРТЫ Использование банка данных является первым необходимым шагом при картографическом анализе любого гена. Но на всех этапах разработки картографической технологии главной целью оставалось картографирование не отдельных генов, а извлечение из карт отдельных генов общей информации о генофонде. Общепринятым методом обобщения тогда был (и остаётся доныне) анализ главных компонент. Этот статистический метод позволяет в изменчивости множества признаков выделить основные тенденции и представить их в изменчивости немногих новых условных признаков — главных компонент. Это метод чисто статистический, но чтобы сделать его картографическим, нужно, казалось бы, совсем немного - рассчитать значения главных компонент для каждой популяции и нанести их на карту. Сложность заключается в том, что разные гены изучены в разных популяциях, а для расчёта главных компонент необходимы данные о значении каждого гена в каждой популяции. Решить эту проблему можно было лишь с помощью карт, основанных на цифровых матрицах. Ведь для таких карт мы имеем значение признака в каждой точке карты. А для другого признака - опять-таки знаем его значение в каждой из тех же самых точек. Таким образом, расчёт главных компонент проводится не по исходным данным, а по картам. Точнее, по картографированным (интерполированным) значениям в каждой точке карты (каждом узле регулярной сетки карты). Карты главных компонент строились и технологией «чернильных пятен». Но когда была создана технология картографирования на основе средневзвешенной интерполяции, и был разработан алгоритм проведения с цифровой матрицей математических операций, то создание карт главных компонент в отечественной геногеографической школе стало обычной и широко используемой процедурой. Каково же было удивление, когда на этом этапе обнаружилось, что в зарубежных статьях (к тому же не в самых последних, а давностью в несколько лет) уже опубликованы карты главных компонент генофонда! Впрочем, изумление перед этим фактом быстро сменилось сознанием того, что такое совпадение подтверждает правильность избранного пути, а также ту огромную востребованность в геногеографии общих параметров генофонда, которая назрела в мировой науке. Очевидно, именно такой была общая логика развития популяционной генетики в 70е и 80е годы, как за рубежом, так и в нашей стране: 1) необходимо обобщить данные о многих генах; 2) был общепринятый метод обобщения - главные компоненты; 3) для содержательной интерпретации результатов нужно было знать географию главных компонент, то есть построить для них карты; 4) для расчёта необходимы данные по каждому признаку в каждой точке карты; 5) получить такие данные возможно только при картографировании каждого признака, причём самым главным результатом картографирования должна быть не сама графическая карта, а лежащая в её основе матрица точных значений признака в каждом узле карты. Самое забавное, что вместо разочарования и глубокого огорчения, что приоритет синтетических карт остался закреплён не за нами, основным чувством была радость - нам стало намного проще объяснять, сколь необходимы геногеография и компьютерные картографические технологии. Если до этого солидные члены солидного Ученого совета заявляли, что подобные нашим контурные карты они рисовали в пятом классе, и нечего заниматься такими пустяками в Академии наук, то теперь авторитет «заграничных» исследований и популярность синтетических карт в мировой науке позволила нам развернуть куда более обширную работу и у нас в России. CAVALLI-SFORZA И ЕГО ШКОЛА Публикации карт главных компонент, о которых мы только что рассказали, принадлежали коллективу, который возглавлял Луиджи Лука Кавалли-Сфорца. Сразу оговоримся, что с историей зарубежной геногеографии авторы знакомы существенно меньше, чем с историей отечественной науки. Поэтому очертим её лишь несколькими штрихами и назовём только два имени. Первое из них принадлежит А. Е. Mourent, организатору и автору крупнейшей сводки данных о частотах генов в населении мира [Mourent et al., 1976]. Этот прекрасный труд сыграл огромную роль для развития геногеографии. На страницах этой книги были сведены данные, кропотливо собранные из сотен публикаций разных лет, причём для каждой изученной популяции приводились подробные сведения о её географическом положении, отнесении к основным регионам мира, характере выборке (пациенты, здоровые, военнослужащие, коренное или «смешанное» население), а также подробная библиографическая ссылка на исходную публикацию этих данных. Для самых изученных генов были приведены и карты их распространения в мире. По сути, этот труд явился прообразом банка данных о генофонде популяций всего мира, а по полноте и объёму данных по классическим маркёрам он не утратил своего значения и сейчас. Второе имя хорошо известно каждому, кто хотя бы бегло знаком с работами по популяционной генетике человека. Лаборатория и научная школа L. L. Cavalli-Sforza являются, пожалуй, наиболее известными в мировой науке. Этим коллективом выполнен целый ряд ставших классическими работ: по совершенствованию методов расчёта генетических расстояний, построению кластеров родства популяций, анализу фамилий как аналога генетических маркёров и множество иных исследований. Именно перу Кавалли-Сфорца принадлежит крупнейшая обобщающая монография в области популяционной генетики человека [Cavalli-Sforza et al., 1994]. Для нашего рассказа наиболее важно, что именно этим коллективом была независимо разработана картографическая технология, в общих чертах аналогичная технологии, созданной отечественной геногеографической школой. Именно эта технология легла в основу названного обобщающего труда, в котором не только приведены геногеографические карты для регионов всего мира, но и дано тщательное описание их генофондов. Главным и важнейшим инструментом для школы Кавалли-Сфорца являются карты главных компонент («синтетические карты» в его терминологии). Каждая из карт интерпретируется как отражение одного из исторических сценариев, сформировавших генофонд популяции, причём предполагается, что более значимые компоненты описывают хронологически более ранние этапы формирования генофонда. СОВРЕМЕННАЯ ОТЕЧЕСТВЕННАЯ ГЕНОГЕОГРАФИЯ В своих важнейших чертах отечественная геногеографическая школа перекликается с только что описанной зарубежной. Это, во-первых, нацеленность на изучение не частной геногеографии отдельных генов, а общих черт строения генофонда. Во-вторых, это широкое использование компьютерных карт, и в первую очередь обобщённых карт (например, главных компонент). В-третьих, применение всего комплекса остальных (не картографических) популяционно-генетических методов для целей геногеографии - выявления объективных закономерностей в структуре генофонда. Особенностью же отечественной школы является в первую очередь наличие развитой и широко разветвлённой картографической технологии [Балановская и др., 1994а, 19946, 1995, 1997]. В разделе 3 Приложения описывается, сколь большой комплекс методов и математических преобразований может быть выполнен с картами отдельных генов. Поэтому, например, результатом работы отечественной школы стало создание не одного, а трех типов синтетических карт: наряду с картами главных компонент создаются и анализируются карты гетерозиготности и карты генетических расстояний. Важным достоинством отечественной картографической технологии является также возможность расчёта корреляций между картами разных признаков, построение корреляционных карт и карт межпопуляционного разнообразия. Одним из последних и самых важных достижений отечественной геногеографии авторы считают внедрение в технологию карт надёжности. Эти карты показывают степень достоверности картографического прогноза на разных территориях в зависимости от обеспеченности исходной информацией, позволяют включать в компьютерные карты лишь данные с высоким уровнем надёжности и исключать из картографирования мало надёжные территории [Нурбаев, Балановская, 1998]. Еще одной особенностью отечественной школы можно считать анализ не только исторической (селективно-нейтральной) изменчивости генофонда, но и выявление действия естественного отбора на конкретные гены. Выше рассказывалось об анализе межпопуляционной изменчивости в генофондах разных регионов и выявленном при этом феномене эквидистантности. Такие устойчивые показатели межпопуляционной изменчивости получаются при усреднении величин изменчивости, характерных для большого числа изученных генов. При этом изменчивость каждого отдельного гена может отличаться от среднего, селективно-нейтрального уровня. И единственной причиной, которая может вызвать достоверные отклонения изменчивости данного гена от средней, является действие естественного отбора на этот ген. Если изменчивость данного гена снижена, можно предполагать действие стабилизирующего отбора. Указанием же на дифференцирующий отбор является ситуация, когда изменчивость рассматриваемого гена оказывается достоверно выше средней изменчивости всей совокупности изученных генов. Именно на этом основана технология выявлении эффектов отбора, опубликованная в работах [Рычков, Балановская, 1990а; Балановская, Нурбаев, 1997, 1998а, 19986, 1998в, 1999]. Итак, мы вкратце описали основные вехи развития геногеографии - *от работ А. С. Серебровского до формирования современной геногеографической технологии, позволяющей проводить всесторонний анализ пространственной изменчивости как отдельных генов, так и генофондов. Подводя общий итог, можно выделить три основные группы методов современной геногеографии: 1) анализ межпопуляционной изменчивости при иерархической организации изучаемых популяций (анализ средней FST); 2) анализ селективной структуры (выявление действия естественного отбора на отдельные гены по отклонениям от средней FST); 3) картографические методы (анализ пространственной структуры генофонда с помощью интерполяционных карт). РАЗНЫЕ ГЕНОФОНДЫ. РАЗНЫЕ ПРИЗНАКИ Рассказывая об истории нашей науки, мы говорили больше об истории идей - истории разработки тех или иных геногеографических методов. И это оправдано, потому что именно наличие мощного метода является главнейшим условием для надёжных исследований конкретных генофондов. Более того, именно разработка методов всегда была стимулом для разработки теоретических вопросов геногеографии. Но неверно думать, что её история сводится лишь к разработке методов. Намного более многочисленны - столь многочисленны, что здесь невозможно перечислить даже главнейшие из них - были исследования конкретных генофондов или конкретных генов. Чтобы дать хотя бы общее представление о масштабе и количестве выполненных геногеографических работ, перечислим некоторые из изученных нами генофондов. Из геногеографически изученных генофондов отдельных народов следует назвать адыгов, башкир, белорусов, марийцев, монголов, осетин, русских, украинцев. Из региональных генофондов - Кавказ, Восточную Европу, всю Европу, Уральский регион, Сибирь, Северную Евразию, Старый Свет (Евразия и Северная Африка). Также изучались генофонды всех основных регионов мира (см. §4 этого раздела). Разнообразие признаков, изучаемых геногеографией, ещё ярче, чем разнообразие изученных ею генофондов. Если изначально геногеография мыслилась как наука о географии генов, то сейчас перечень используемых для описания генофонда маркёров намного шире. Это, во-первых, антропологические признаки (соматология, дерматоглифика, одонтология). Во-вторых, данные археологии. Одно из важнейших геногеографических исследований - генофонда палеолита - выполнено не на данных о частотах генов, а на данных о признаках материальной культуры (орудия, искусство, жилища) и фауны разных этапов палеолитической эпохи. В-третьих, в современной геногеографии широко используется анализ квазигенетических маркёров, в первую очередь фамилий. Время от времени геногеографические технологии применяются и для признаков, вовсе далёких от генетики, таких как заболеваемость, техногенная напряжённость среды, гормональный статус или показатели антропологического развития детей. Правда, в этих случаях решаются не столько популяционные, геногеографические задачи, сколько геногеографические технологии используются для решения задач смежных наук. Но конечно, большинство геногеографических исследований проводились на основе данных о частотах генетических маркёров, причём всех их типов: и классических, и аутосомных гипервариабельных ДНК маркёров, и инсерционно-делеционного ДНК полиморфизма, и маркёров митохондриальной ДНК, и гаплогрупп Y хромосомы. 2Выбор генохронологически датируемых событий, равно как и их описание, были осуществлены профессором Ю. Г. ычковым и приводятся по работе [Балановская, Рычков, 1990 а]. Все работы проведены в 60е-80е годы его сотрудниками и выполнены под его руководством [Рычков, 1984, 1980]. 3Вопросы стационарности генетического процесса мы подробно обсуждали в работе [Балановская, Нурбаев, 1997]. 4Даже если сейчас данный аллель селективно-нейтрален - он мог быть адаптивно значимым на прошлых временных срезах, на иных поворотах истории генофонда. При анализе селективной структуры генофонда мира по всем наиболее изученным классическим маркёрам, мы, к своему удивлению, не обнаружили ни одного аллеля, который бы во всех регионах мира и на всех этапах истории был бы селективно-нейтральным! [Рычков, Балановская. 1990 б; Балановская, Рычков, 1997]. Более того, аллели во времени и в пространстве могут не только переходить из селективно-нейтральных в разряд селективно-значимых и наоборот - может меняться даже вектор отбора! То есть один и тот же вариант гена может быть подвержен то стабилизирующему, то дифференцирующему отбору на разных отрезках времени и пространства мирового генофонда. 5 Мы здесь приводим для каждого из регионов его реальные наблюдаемые значения FST, полученные нами по классическим маркёрам. В Средней Сибири разнообразие популяций внутри этноса FST(MS-POP)=4.41, различия между этносами FST(MS-ET)=2.59, а общие различия в пределах региона FST(MS)=7.00. В Восточной Сибири разнообразие популяций внутри этноса FST(ES-POP)=4.01, различия между этносами FST(ES-ET)=2.36, а общие различия в пределах региона FST(ES)=6.37. 5 Мы избегаем конкретных ссылок в этом случае. Кто из нас застрахован от таких или иных ошибок? |
загрузка...