6.2. Митохондриальная днк (материнские линии)
|
Самый изученный маркёр - Его особенности
§1. Изученные русские популяции: Кто их изучал - Почему обязательно «исконный» ареал? - Качество выборок - Семь «хороших» популяций §2. География гаплогрупп: Самая частая гаплогруппа - Самые редкие гаплогруппы - Умеренные гаплогруппы - Особый профиль Русского Севера - Редкость «азиатских» гаплогрупп у русских - Пенеги - Древние пласты Севера? §3. Русские популяции в генетическом пространстве мтДНК: Главный кластер популяций - Особость севера - Призрачное своеобразие юга - Значительная дифференциация русского генофонда - Выявляемая при корректном анализе §4. Сказка об изолах и миграх: Генетическая память популяции §5. Прокрустово равенство: сколько «русских» гаплотипов у соседей по Европе? Ищем общие гаплотипы - Ненасытная Харибда - Коварная Сцилла - Мужественный Прокруст - Общий восточно-европейский генофонд - Русские и финно-угры - Русские и славяне Я научила женщин говорить... Но, Боже, как их замолчать заставить! Анна Ахматова С девяностых годов XX века и до настоящего времени митохондриальная ДНК (мтДНК) является, пожалуй, наиболее интенсивно изучаемым ДНК маркёром. Одна из причин такого внимания к мтДНК кроется в её малости (в сравнении с ядерным геномом). Это позволило унифицировать набор маркёров мтДНК. Если выбор аутосомных маркёров огромен, и разные лаборатории изучают различные гены, то мтДНК стала общим местом популяционных исследований. Значение этого трудно переоценить - стандартизация программы популяционных исследований [Macaulay et al., 1999] позволила сосредоточить усилия многих и многих лабораторий на одной и той же системе маркёров, и накопить в общемировом масштабе внушительный объём данных о полиморфизме мтДНК (глава 9). Благодаря концентрации усилий на одной генетической системе, её полиморфизм стал досконально изученным. Это стимулировало и разработку методологических вопросов - анализа медианных сетей, датировок времени возникновения аллелей (гаплогрупп) [Forster et al., 1996; Saillard et al., 2000], сравнения новых данных с результатами работ по классическим маркёрам. Стали обсуждаться вопросы воспроизводимости результатов популяционных исследований при повторном изучении тех же популяций [Yao et al., 2002] и, соответственно, различных стратегий формирования выборок. Многие иные проблемы, ранее едва просматривавшиеся на заднем плане ДНК исследований, встали на повестку дня, как только был накоплен определённый опыт беспрецедентно подробного (по масштабам ДНК исследований) изучения географии митохондриальной изменчивости. Эта наработанная методология анализа нерекомбинирующих генетических систем уже перенесена на анализ Y хромосомы, и, видимо, будет широко использоваться и для гаплотипов других маркёров - X хромосомы и аутосомных. Возможно, такой взгляд на мтДНК - просто как на самый изученный ДНК маркёр - покажется странным на фоне научной моды на использование мтДНК в решении проблем микроэволюции, филогеографии. Однако такой взгляд позволяет сравнивать разные маркёры не только «потенциально» - по их теоретическим возможностям для изучения генофонда, но и «кинетически» - по той энергичности, с которой они изучаются в практике популяционных исследований. Маршрут раздела таков. Сейчас мы вкратце рассмотрим особенности мтДНК - этот раздел читатель может смело пропустить и возвратиться, если последующее изложение покажется непонятным. Затем в §1 мы рассказываем, что к настоящему времени сделано для изучения «митохондриального генофонда» русских популяций. Далее представляем попытку обобщения накопившейся информации - анализа изменчивости мтДНК в пределах русского генофонда и сравнения его с соседними группами населения на уровне гаплогрупп (§2-§3) и на уровне гаплотипов (§4-§5). ОСОБЕННОСТИ МИТОХОНДРИАЛЬНОЙ ДНК ДВА СВОЙСТВА. Для изучения генофонда важны две особенности митохондриальной ДНК. Во-первых, мтДНК наследуется только по материнской линии: каждый человек получает свой гаплотип мтДНК от матери, то есть в передаче мтДНК принимают участие только женщины. Популяционному генетику это даёт возможность прослеживать популяционные процессы (миграций, динамики численности и т.д.), в различной степени затрагивающие представителей разных полов. Во-вторых, у мтДНК нет рекомбинации, т. е. нет «перетасовывания» генов, пришедших от матери и пришедших от отца (просто потому, что от отца мтДНК не приходит). Благодаря этому вся мтДНК передаётся в поколениях единым «текстом» - гаплотипом. Это позволяет вычленять «линии» мтДНК (похожие «тексты», связанные единством происхождения), выявлять их «родословные», датировать время расхождения основных ветвей таких родословных, объективно фиксировать события возникновения новых вариантов мтДНК и пути их передвижения по Земле вместе с потоками мигрантов. Эти свойства вызвали всеобщий интерес к мтДНК - к настоящему времени на территории Европы почти не осталось народов, для которых не имелись бы данные по мтДНК, причём многие получены по обширным выборкам. УСТРОЙСТВО мтДНК. Митохондриальная ДНК представляет собой небольшую молекулу, расположенную в клетке особняком - не в ядре, как основная часть генома, а в митохондриях. У мтДНК есть свои особенности метаболизма. Так, например, считается, что мутации в митохондриальной ДНК происходят примерно в 100 раз чаще, чем в ядерной. МтДНК содержит немногим более 16000 нуклеотидов («букв», которыми написан текст мтДНК). Каждому из них присвоен номер, определяющий его место - позицию - в тексте. В митохондриальной ДНК можно выделить два типа участков. Первый - участки, кодирующие митохондриальные белки; они составляют основную часть (более 15000 нуклеотидов) мтДНК. В этих участках мутации редки - то есть случайные ошибки при передаче текста, когда какая-то «буква» вдруг заменяется другой, встречаются редко, поскольку отсеиваются отбором. Однако известно довольно много позиций (определённых мест в пронумерованной последовательности нуклеотидов мтДНК), в которых у разных людей может находиться не строго определённый, а тот или иной нуклеотид. В основном они находятся в некодируюшей части мтДНК. Считается, что большинство этих мутаций произошли за историю человечества лишь однажды, и поэтому все люди, имеющие данную мутацию, являются между собой более или менее близкими родственниками по материнской линии. Во всяком случае, они связаны более тесным родством, чем те, у кого нет данной мутации. ГАПЛОГРУППЫ. Нужно оговориться, что повторные мутации в той же позиции изредка все-таки случаются. Поэтому отдельно взятая мутация в мтДНК может и не быть свидетельством общего происхождения, а произойти независимо. Надежное определение родства молекул мтДНК проводится по наличию не одной, а нескольких общих мутаций (как в кодирующей, так и в некодируюшей частях мтДНК). Сочетание мутаций однозначно определяет гаплогруппу - т. е. совокупность («линию») митохондриальных ДНК, имеющих общую предковую форму. Однако индивидуумы, молекулы митохондриальной ДНК которых относятся к одной и той же гаплогруппе (т. е. имеющих одинаковые нуклеотиды в позициях, «диагностирующих» гаплогруппу) могут различаться по другим участкам, в частности, гипервариабельным сегментам. ГАПЛОТИПЫ. Два таких участка (гипервариабельный сегмент 1 и гипервариабельный сегмент 2 - ГВС1 и ГВС2) представляют оставшуюся часть митохондриальной ДНК. Эта часть мтДНК не кодирует белки, в отличие от основной «кодирующей части». Каждый из этих сегментов состоит примерно из 400 нуклеотидов. Если при изучении кодирующей части обычно проверяются выборочно только несколько нуклеотидов1, характерных для той или иной гаплогруппы, то при анализе ГВС1 проводится прямое «секвенирование» - определяется, какой именно нуклеотид расположен в каждой из 400 позиций этого сегмента. Иными словами, анализируются не несколько, а все 400 ДНК маркёров ГВС1. Данные по ГВС1 позволяют выделить не несколько гаплогрупп, а огромное множество вариантов мтДНК (гаплотипов). То есть каждую гаплогруппу можно подразделить на субгаплогруппы, а их - на целый ряд гаплотипов. Это многократно повышает разрешающую возможность анализа, в том числе определения возраста гаплогрупп. Скорость накопления мутаций в некодирующих участках намного выше, чем в кодирующей части (поэтому они и называются гипервариабельными), - здесь одни и те же мутации часто возникают независимо, многократно. Поэтому точная классификация типов митохондриальной ДНК (гаплотипов) производится в два этапа: сначала для данной линии определяется принадлежность к той или иной гаплогруппе (на основании изучения кодирующей части), а затем точное систематическое положение внутри гаплогруппы (по ГВС1). Обычно кодирующую часть «смотрят» лишь на несколько ключевых мутаций, а для определения субгаплогруппы и гаплотипа привлекаются данные по ГВС1. ФИЛОГЕОГРАФИЯ. Зная достаточное количество сочетаний мутаций в ГВС1, встреченных в популяции, можно с большой долей вероятности определить порядок возникновения этих мутаций во времени и, соответственно, составить «родословное древо» для гаплотипов митохондриальной ДНК. Это древо обычно представляется в виде так называемой медианной сети, которая отличается от обычной дендрограммы тем, что отдельные ветви могут сливаться друг с другом. Такое слияние ветвей указывает, что нам пока неизвестно, шло ли развитие по одному, другому, или обоим направлениям, но наличие какого-либо третьего пути маловероятно. Установление такой филогенетической классификации митохондриальной ДНК имеет большое значение для популяционных работ, поскольку позволяет оценить число произошедших мутационных событий и, при ряде допущений об их скорости, вычислить время, прошедшее с момента возникновения данной гаплогруппы (ветви родословного древа мтДНК). Подобно тому, как знание времени чеканки монеты, обнаруженной в археологическом памятнике, может помочь датировать памятник, так и знание времени возникновения гаплогруппы, обнаруженной в популяции, может способствовать лучшему пониманию истории популяции. В обоих случаях, мы не знаем точной датировки популяции или археологического памятника (ведь монета или гаплогруппа могли бережно храниться и передаваться в долгой чреде поколений, с незапамятных времен). Но мы точно знаем, что памятник возник позднее, чем время, выбитое на монете. Поэтому, когда мы слышим, что датировки мтДНК восточных славян составляют 30 тысяч лет, не стоит гордиться, что славяне уже 30 тысяч лет существуют на свете. Это лишь означает, что славяне появились позднее, чем 30 тысяч лет назад (а когда именно - по-прежнему неизвестно). Насколько ценна такая информация, можно спорить. Но если с ней умело обращаться, то она может помочь установить, что одни древние популяции древнее других. Изучив мтДНК, можно с высокой достоверностью определить эволюционные соотношения всех гаплотипов - как представленных в современном генофонде, так и отсутствующие «промежуточные формы». Это позволяет реконструировать процесс возникновения гаплотипов, локализовать во времени и в пространстве происхождение каждой конкретной гаплогруппы мтДНК. Эта область исследований получила название филогеографии. Сопоставление современного ареала гаплогруппы с древней зоной её возникновения открывает путь к прослеживанию миграций населения, а также иных событий демографической и этнической истории (сокращения численности и «экспансии», подразделение популяции, особенностей брачной структуры и других). ДВА МАСШТАБА АНАЛИЗА. Изменчивость «митохондриального генофонда» можно изучать в двух «масштабах». Первый - масштаб гаплогрупп. Обычно рассматривают распределение гаплогрупп по территории, сравнивают частоты гаплогрупп в разных популяциях, в том числе методами многомерного анализа: главных компонент, многомерного шкалирования, кластерного анализа. Второй - масштаб гаплотипов. Он позволяет проводить более тонкий анализ. Здесь можно определить уровень гаплотипического разнообразия, сравнить спектр (набор) гаплотипов, встреченных в разных популяциях. Своеобразный промежуточный уровень - разнообразие гаплотипов в пределах конкретной гаплогруппы. Зная величину разнообразия гаплотипов, можно рассчитать время, за которое это разнообразие могло накопиться в ходе мутационного процесса, то есть время, прошедшее с момента возникновения гаплогруппы. В §2 и §.3 мы проведём анализ в масштабе гаплогрупп, а затем перейдём к масштабу гаплотипов (§4 и §5). §1. Изученные русские популяцииРусский народ - самый многочисленный в Европе и занимает самый большой ареал. Поэтому от его митохондриального генофонда можно ожидать значительного разнообразия. Мы видели (главы 2, 4), что вклад тех ли иных антропологических пластов различен в разных регионах. Поэтому данные по генетике «одиноких» русских популяций отражают лишь один из фрагментов генофонда и не могут дать характеристику русского народа в целом. А значит, необходимо изучить полиморфизм мтДНК во многих регионах, на всей обширной территории «исконного» ареала русского народа. ИСТОРИЯ ИЗУЧЕНИЯ Самые первые данные о мтДНК в русских популяциях появились давно [Лемза, Соколова, 1992; Петрищев. Кутуева, 1993; Казаковцева и др, 1998], но из-за методических различий (тогда типировались лишь немногие ПДРФ маркёры) они мало приложимы к современным исследованиям. В работе [Orekhov et al, 1999] впервые были представлены данные о полиморфизме гипервариабельного сегмента мтДНК у русских - суммарно 103 человека из Костромской, Курской и Рязанской областей. К сожалению, в статье не указано, к какой из популяций относится каждый гаплотип. Поэтому сейчас, когда имеются данные по многим региональным популяциям, эти самые первые данные, как ни грустно, приходится оставлять за бортом нашего анализа генофонда. Для нас это яркий пример того, насколько важно при начале работы уделять внимание формированию выборок, а при её завершении - возможно полной публикации исходных данных и популяционных сведений о них. Несомненно, что при своем появлении, на фоне практически полного отсутствия данных по российским популяциям, эта работа была своевременной. Но если бы для каждого из гаплотипов была дана «привязка» к географически конкретным популяциям и по каждой популяции выборка была бы достаточна, то эта статья не потеряла бы своего значения и сейчас. К настоящему времени в работах Б. А. Малярчука опубликованы данные по шести2 русским популяциям со средним размером выборки N=66 [Малярчук и др, 2001, 2002]. В этих статьях представлены результаты ПДРФ анализа кодирующей части мтДНК для всех шести популяций, тогда как данные по сиквенсу ГВС1 опубликованы в отдельной статье для трёх из этих популяций. Соответственно, для трёх других популяций гаплогруппы определены с тем уровнем разрешения, который легко достигается с помощью небольшого числа ПДРФ маркёров. Этот уровень заметно ниже того, который достигается при комбинации информации ПДРФ и ГВС1. Отсутствие подразделения гаплогруппы U на субгаплогруппы в этих статьях особенно затрудняет их использование. Данные по трём из этих популяций опубликованы также в [Malyarchuk et al, 2002] где проведён анализ как ПДРФ, так и ГВС1. Правда, в статье не была указана принадлежность конкретных гаплотипов к той или иной популяции. Авторы благодарны Б. А. Малярчуку, предоставившему им такую информацию. Она позволила определить гаплогруппы с общепринятым уровнем разрешения, и эти данные использованы в §5 для графика многомерного шкалирования. Также имеются данные по русскому населению Ростовской области [Richards et al, 2000; Генофонд и геногеография...Т.2, 2003]. Северная популяция (Каргополь, N=76) изучена в работе [Belyaeva et al, 2003] и по ГВС1, и по ПДРФ кодирующей части. Авторы руководили сбором экспедиционных образцов в этой популяции, поэтому на эту выборку мы полагаемся уверенно. Авторы этой книги, совместно с Эстонским биоцентром, возглавляемым Р. Виллемсом, также предприняли изучение полиморфизма мтДНК в пяти русских популяциях (средний объём выборки N=130). Все экспедиции были проведены коллективом под руководством Е. В. Балановской. Обследование северовосточной (Архангельская область) и восточной (Костромская область) популяций проведено авторами книги. При работе в западной популяции (Смоленская область) нам помогали А. С. Пшеничное и Н. Н. Аболмасов. Обследование южных популяций проведено нашими коллегами из этих регионов: кубанских казаков - Э. А. Почешховой, а в Белгородской области сотрудниками М. И. Чурносова. Все экспедиции проведены по единому плану и с соблюдением строгих требований к формированию выборки. Все генотип и рование мтДНК проведено О. П. Балановским на базе Эстонского биоцентра. Изучен как полиморфизм ГВС1, так и ПДРФ анализ кодирующей части мтДНК. Эти данные будут опубликованы в ближайшее время, и анализируются в данной книге. В литературе имеются ещё данные по русскому населению Магадана, Башкирии, Томска и Новосибирска. Однако эти популяции находятся далеко за пределами «исконного» русского ареала, и потому здесь не рассматриваются. ДАЛЬНЕВОСТОЧНЫЕ СЛАВЯНЕ Конечно, такой подход изучения - только коренного населения на его «исконном» ареале - хотя и служит краеугольным камнем нашей книги, но не бесспорен. Можно предположить, что популяция «дальневосточных славян» (русские и украинцы Магадана, как пишут изучавшие их авторы, «происходящие из различных регионов бывшего СССР» [Malyarchuk, Derenko, 2001]) представляет не меньший интерес, чем популяции восточных славян. Действительно, «неинтересность» дальневосточных славян объясняется лишь тем, что эта популяция сформировалась всего несколько десятков лет назад и представляет собой сборную группу людей, происходящих из самых различных частей СССР. Но ведь тысячу лет назад восточнославянское население Волго-Окского междуречья также, по всей видимости, являлось чрезвычайно молодым образованием, сотканным из переселенцев из самых различных, более западных областей Киевской Руси. Почему же тогда мы считаем генофонд восточных славян интереснейшим объектом, а дальневосточных славян оставляем за бортом исследования? Неужели дело лишь в очаровании древности? На этот вопрос не так просто ответить - особенно потому, что перед очарованием древности, и вправду, нам устоять нелегко. Но дело все же в другом. Сейчас, тысячу лет спустя, мы знаем, что восточные славяне Волго-Окского междуречья оставили многочисленное потомство, образовали устойчивые популяции и стали ядром русского народа, существующего уже в течение многих веков и широко расселившегося - вплоть до Дальнего Востока. А вот какова дальнейшая судьба дальневосточных славян? Сформируется ли на их основе устойчивая в поколениях популяция? Или уже через несколько лет в тех местах будут жить новые мигранты, а нынешнее население, или их дети, будут населять другие города и страны? Неизвестно. Но очень велика вероятность второго исхода! А в таком случае генетически изученная популяция уже через несколько лет после её изучения перестаёт существовать, и опубликованные данные перестают соотноситься с каким-либо генофондом. Напротив, данные по устойчивой популяции продолжают адекватно описывать её генофонд и по прошествии нескольких поколений. Именно поэтому (этот вопрос рассматривается в главе 1 ив Приложении) мы изучаем лишь «истинные» популяции, одним из критериев которых является их устойчивость в чреде поколений и заключение не менее 50% браков внутри популяции. И именно поэтому каждый народ изучается на его «исконной» территории - там, где в течение веков формировался генетический портрет этого народа, какая бы судьба его не постигла далее (он колонизировал или его колонизировали). ВЫБОРКИ В популяциях, обследованных нашим коллективом, мы всегда стремились к тому, чтобы размер выборки составил около 150 человек - в два раза больше, чем принятый для анализа аутосомных маркёров. Увеличение выборки необходимо, чтобы компенсировать гаплоидность мтДНК3 и возможно более полно охватить разнообразие гаплотипов4. Обследовались группы деревень или небольшие города вдали от магистралей - с целью избежать влияния недавних миграций. Письменное информированное согласие было получено от каждого из обследованных. Все четверо бабушек и дедушек обследованных были рождены в пределах ареала данной популяции. На практике это означает, что у подавляющего большинства обследованных все их предки происходили из того административного района данной области, в котором проводилось экспедиционное изучение. Остальные генеалогические линии обследованных ведут к соседним районам той же области. Таким образом, благодаря тщательному анкетированию обследованных людей, собранные выборки действительно репрезентативно представляют коренное население изученных регионов. Аналогично собрана и выборка русских Каргополя - оба автора принимали деятельное участие в работе этой экспедиции. Что же касается популяций, обследованных Б. А. Малярчуком, то требования при формировании выборок были менее строгими: обследование проводилось на базе областных больниц и станций переливания крови, изучалось население крупных городов, и основное внимание было уделено русской национальности и месту рождения обследованных и их матерей [Малярчук, Деренко, 2004]. То есть анкетирование проводилось до второго, а не третьего поколения, как в наших выборках. И единицей обследования выступала область, а не район (меньшая географическая точность). Эти методические различия могут иметь важное следствие: результаты недавних (происходивших в XX веке) миграций могут намного сильнее проявляться в данных Б. А. Малярчука. Тем самым эти данные могут в большей мере представлять смешанное (а не коренное) русское население, проживающее в изученных регионах. Это необходимо иметь в виду при интерпретации практически всех результатов, поэтому при каждом виде анализа мы оговариваем, данные по каким именно выборкам были использованы. В §3 мы ещё раз вернемся к этому вопросу. АНАЛИЗИРУЕМЫЕ ПОПУЛЯЦИИ В таблицах 6.2.1. и 6.2.2. указаны все выше перечисленные популяции. Можно видеть, что изучены различные региональные группы, которые охватывают практически весь «исконный» русский ареал. Из них семь популяций (Каргополь, Пинега, Унжа, Смоленск, Белгород, кубанские казаки, юго-восточные русские) представлены качественными выборками, тогда как данные по остальным популяциям, как только что описывалось, по тем или иным параметрам не подходят для геногеографического анализа. Поэтому основной анализ выполнен по семи перечисленным популяциям. Это число популяций, к сожалению, недостаточно для проведения картографического анализа. Но оно позволяет набросать митохондриальный портрет русского населения, выявить особенности региональных популяций, оценить межпопуляционную изменчивость в пределах русского генофонда. §2. География гаплогруппАнализ изменчивости и антропологических, и классических генетических маркёров мы начинали с «простых» карт отдельных признаков (главы 4, 5). Но для мтДНК количество изученных популяций пока недостаточно для картографирования. Слишком схематичны и неточны были бы карты, построенные всего по семи популяциям. Поэтому мы решили попробовать пока без карт уловить тенденции в географическом распределении гаплогрупп. И основной тенденцией вновь оказалась широтная изменчивость, которая столь ярко проявилась при анализе классических маркёров и антропологических признаков (главы 4, 5): деление русского ареала на север и юг. ГАПЛОГРУППА Н Частота этой «европейской» гаплогруппы варьирует в русских популяциях в широких пределах. Она резко падает с севера на юг: от почти 50% в Архангельской области до 35-40% у южных русских, при промежуточных частотах в центре ареала. Снижение частоты гаплогруппы Н к югу характерно и для населения Европы в целом. ГАПЛОГРУППЫ I, Ul, U2, U3, X Эти гаплогруппы встречены только в южной половине русского ареала, причём практически в каждой изученной популяции, но при этом отсутствуют в северных популяциях (Пинега, Каргополь). Конечно, это отсутствие временное - при дальнейшем увеличении выборок эти гаплогруппы скорее всего обнаружатся и на севере, но можно полагать, что их частоты будут намного ниже, чем на юге. Все эти пять гаплогрупп редкие: средняя частота у русских 1-2%. ГАПЛОГРУППЫ К, R, Т, U5A, U5B Эти гаплогруппы более частые и встречены повсеместно. Но и для них частоты на севере и на юге различны: гаплогруппы К, R и U5b встречаются на Русском Севере несколько чаще, а Т и U5a - реже, чем в остальных русских популяциях. ВОСТОЧНО-ЕВРАЗИЙСКИЕ ГАПЛОГРУППЫ А, С, Р, Z Любопытно распределение восточно-евразийских гаплогрупп. Этот термин обозначает гаплогруппы, основная зона распространения которых - Восточная и Центральная Азия (раздел 9.2.). Поэтому восточно-евразийские гаплогруппы могли бы маркировать монголоидный вклад в генофонд. Когда мы рассматриваем не только русские, а все популяции Восточной Европы, то география восточно-евразийских гаплогрупп очень четкая - каждая из гаплогрупп чаще встречается на востоке региона (глава 8). Что же касается русских популяций, то такого тренда мы у них не видим: восточно-евразийские гаплогруппы встречаются в них хотя и с низкой частотой, но повсеместно. Уже не первый раз в этой книге мы сталкиваемся с тем, что распределение генов в русском ареале отнюдь не следует закономерностям, которые выявлены в глобальном масштабе. Суммарная частота всех восточно-евразийских гаплогрупп в русских популяциях (2.2%) практически так же низка, как и в населении Западной Европы (например, в популяциях французов в среднем 0.5%). При этом в популяциях Восточной Азии эти гаплогруппы доминируют, составляя 85-100% от всех гаплогрупп у монголов, китайцев, эскимосов. Таким образом, данные по митохондриальной ДНК указывают на отсутствие сколько-нибудь значительного монголоидного пласта в русском генофонде. Впрочем, хотя для отдельных восточно-евразийских гаплогрупп в русском ареале тренды не обнаруживаются, их суммарная частота может оказаться информативным показателем. Хотя она для русских и невелика (2.2%), но прослеживается тенденция её некоторого увеличения к востоку. Так, в восточной русской популяции (Унжа) суммарная частота восточно-евразийских гаплогрупп - 3.8%. На северо-востоке русского ареала (Пинега) суммарная частота ещё чуть выше - 6.2%, правда, она возрастает только за счет одного очень специфичного варианта гаплогруппы D, частота которого достигает 5.6% и который связан скорее с севером Европы, чем с Азией (см. ниже «Пенеги»), Так что высокая доля восточно-евразийских гаплогрупп в этих двух восточных русских популяциях может быть связана с особенностями самого древнего пласта генофонда на Русском Севере. К тому же выборка Унжи невелика. Так что только дополнительные детальные исследования смогут показать, есть ли хоть сколько-нибудь реальное повышение частот восточно-евразийских гаплогрупп у восточных русских. ПЕНЕГИ Эта пинежская популяция вообще наиболее своеобразна: здесь не обнаружены гаплогруппы V, X и U5a, тогда как U5b присутствует с частотой в два раза выше, чем в других русских популяциях. Последняя черта могла бы сближать эту популяцию с саамами (генофонд которых представлен в основном лишь двумя гаплогруппами - U5b и V), но «саамская» гаплогруппа V отсутствует в популяции пенегов. Северно-русская популяция в верховьях р. Пинеги обследована нами в 2001 г. Экспедиция была проведена как комплексная: она включала и сбор образцов крови для анализа ДНК и классических маркёров; и сбор генетико-демографической информации; и сбор данных о квазигенетических маркёрах (фамилии); и антропологическое обследование (расовая программа и антропологическая фотография). Антропологический раздел осуществляли акад. Т. И. Алексеева и к.б.н. А. Г. Новожилов. Освоение Русского Севера является одной из вех в этногенезе русского народа. Особое внимание привлекают те группы, где предполагается значительное влияние субстратного дославянского населения. К ним относится и данная популяция, расположенная на северо-востоке Архангельской области на самой границе с республикой Коми. Нами был обследован ряд сельсоветов Пинежского района - Нюхченский, Сосновский, Сурский, Ново-Лавелевский - тянущиеся лентой вдоль Пинеги. Результаты антропологического анализа показали, что население данной популяции близко не к субстратному населению восточных финно-угров, а к старожильческому русскому населению: выявленный антропологический тип полностью соответствует размаху изменчивости северного русского типа [Алексеева и др, в печати]. Население является высокорослым, с ярко выраженными чертами европеоидного типа, выраженной горизонтальной профилировкой лица, повышенным процентом светло-голубых глаз (№ 10-11) и русых волос, сильно выступающим носом. В целом, этот тип отражает влияние не низовской (Ростово-Суздальской) колонизации Русского Севера, а влияние более ранней северо-западной (Новгородской) русской колонизации и. возможно, балтского компонента. Но не исключено и влияние западных финно-угров, расселившихся на Пинеге до прихода сюда и славян, и коми («Сура поганая» русских источников). Итак, по антропологическим данным пинежская популяция сближается с более западными популяциями. Что же показывает мтДНК? В таблице 6.2.3. приведены генетические расстояния от пинежской популяции до тех популяций Европы, с которыми могло быть связано её происхождение. 1) Малые генетические расстояния (менее 0.05). Наиболее сходным население Пинеги оказывается с другой северной русской популяцией (Каргополь). Также пинежские русские генетически сходны с немцами (!), литовцами и поляками, то есть очерчивается круг популяций, расположенных намного западнее, чем сама Пинега. 2) Средние генетические расстояния (0.05-0.07). Популяции, среднеудаленные от Пинеги, охватывают широкий круг славянских популяций (русские, украинцы, чехи), а также одну балтскую (латыши) и одну западную финно-угорскую популяцию (эстонцы). 3) Большие генетические расстояния (более 0.07). Популяции, генетически значительно отличающиеся от пинежских русских - в основном финно-угорские (карелы, финны, коми). Особенно удивляет отличие пинежской популяции от соседних коми. В момент обследования население по Пинеге имело столь тесные контакты с республикой Коми, что предпочитало работать вахтовым методом там, а не в собственном районном центре (Карпогорах). Этнографические, культурные заимствования от коми в этих русских популяциях велики [Жеребцов, 1982], что подтверждает реальность долговременного контакта с коми. Но при этом по частотам гаплогрупп мтДНК пинежская популяция резко отличается от коми и вообще от финно-угорских популяций, и сближается с широким кругом балто-славянских популяций. Таким образом, данные по мтДНК свидетельствуют, что пинежская популяция в целом генетически не связана с финно-угорским субстратом. Такой результат стал для авторов неожиданностью, поскольку вся совокупность данных, рассматриваемых в этой книге, свидетельствует как раз о значительной или даже определяющей роли финно-угорского субстрата в формировании структуры русского генофонда. Данные по пинежской популяции (где, как мы видим, этот субстрат не выявляется) свидетельствуют, что эта популяция не укладывается в общую картину. А это, в свою очередь, говорит, что наша картина является слишком упрощённой, и нужны бы специальные усилия по приведению её в большее соответствие с ходом реального, сложного процесса смешения славянских и финно-угорских популяций. Тем любопытнее, что те же данные по мтДНК, которые (в целом) показывают различия пинежской популяции и финно-угров, показывают их сходство в одной характерной частности. Речь идет о любопытном варианте гаплогруппы D с мотивом 126-136, который представлен на Пинеге двумя гаплотипами. Именно этот мотив был обнаружен и у саамов [Tambets et al., 2004]. Вообще, этот вариант гаплогруппы D географически приурочен к северу Восточной Европы - от финнов до северо-восточных русских и коми. Вполне возможно этот вариант маркирует древнюю общность генофонда на этой обширной территории. 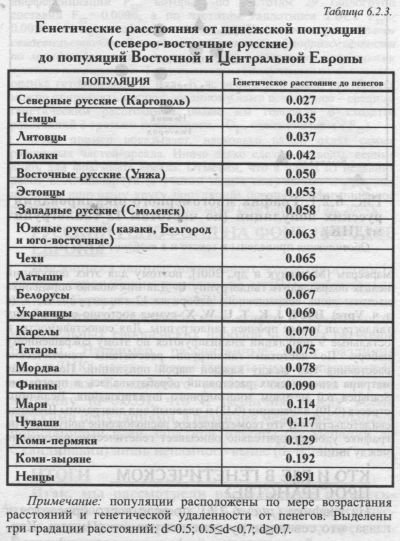
Генетические расстояния от пинежской популяции (северо-восточные русские) до популяций Восточной и Центральной Европы §3. Русские популяции в генетическом пространстве мтДНК Мы рассмотрели изменчивость мтДНК русских популяций в географическом пространстве. Теперь рассмотрим положение русских популяций в генетическом пространстве. Наиболее полную картину даёт график многомерного шкалирования, показывающий генетические взаимоотношения популяций (рис. 6.2.1). МЕТОД МНОГОМЕРНОГО ШКАЛИРОВАНИЯ По частотам всех основных гаплогрупп рассчитаны генетические расстояния между всеми изученными русскими популяциями. Эту матрицу генетических расстояний мы уже использовали при сравнении пинежской популяции с соседями. В матрице содержится информация о генетическом сходстве между всеми возможными парами популяций. Метод многомерного шкалирования позволяет изобразить это сходство на графике. Каждая популяция изображается в виде точки. И расстояния между каждой парой точек соответствуют генетическому сходству между этой парой популяций. Чем больше популяций, тем сложнее изобразить их взаимное расположение на плоском графике. Метод многомерного шкалирования как раз и позволяет создать такой график с минимальным искажением. График основан на частотах 12 гаплогрупп и показывает взаимное генетическое сходство 12 русских популяций (перечислены в табл. 6.2.1). Три из этих популяций (Краснодарская, Белгородская и Нижегородская) изучены только по ПДРФ маркёрам [Малярчук и др, 2001], поэтому для этих популяций нельзя подразделить гаплогруппу U. Для них можно определить частоты лишь сокращенного набора из 12 гаплогрупп: Н, V (в т. ч. Vpre), HV, I, J, К, Т, U, W, X, сумма восточно-евразийских гаплогрупп и «все прочие» гаплогруппы. Для сопоставимости, и остальные 9 популяций анализируются по этому сокращенному набору. По частотам гаплогрупп рассчитаны генетические расстояния Нея между каждой парой популяций. Полученная матрица генетических расстояний обрабатывалась в программе Statistics 6.0 методом многомерного шкалирования. Величины стресса (0.10), алиенации (0.13) и плавный вид диаграммы Шепарда свидетельствуют, что геометрическое расположение популяций на графике удовлетворительно описывает генетические расстояния между ними. КТО И ГДЕ В ГЕНЕТИЧЕСКОМ ПРОСТРАНСТВЕ? На графике (рис. 6.2.1), прежде всего бросается в глаза, что северные русские популяции (Пинега, Каргополь - обозначены на графике квадратами) размещаются в отдалении от основного кластера, в котором собрались практически все остальные русские популяции (кружки). Тем неожиданнее выглядит поведение самых южных русских популяций (обозначены треугольниками). Одна из них (Кубань) входит в основной кластер, а две прочих (Ставрополь и Краснодар) разлетелись в разные углы графика. Все три популяции (Краснодарского, Ставропольского краев и кубанские казаки Адыгеи, табл. 6.2.1.) представляют русское население Предкавказья, которое обычно рассматривается как единый южнорусский тип. 
Рис. 6.2.1. График многомерного шкалирования русских популяций (по частотам 12 гаплогрупп мтДНК). ОТ ЧЕГО РАЗЛЕТЕЛИСЬ ЮЖНЫЕ ПОПУЛЯЦИИ? Столь разительные отличия между этими популяциями по маркёрам мтДНК, конечно, в какой-то степени могли бы быть вызваны смешением с соседними народами Кавказа, которые различаются между собой по спектру гаплогрупп. Но трудно представить, чтобы генофонд, к примеру, Краснодарского края определялся смешением именно с соседними коренными народами Северного Кавказа - слишком несоизмеримы численности населения. Тем более, что наши данные по аутосомным ДНК маркёрам показывают, что даже у кубанских казаков, издавна проживающих бок о бок с адыгами, не обнаруживается поток генов между этими популяциями! И данные по структуре браков Краснодарского края свидетельствуют, что обычно один из супругов происходит из далекого региона: Краснодарский край вбирает в себя мигрантов со всей России. Можно полагать, что такой тип брачной структуры относится не только к современности, но издавна характерен для русского населения южных регионов России [Наследственные болезни в популяциях.., 2002]. Такой «межрегиональный» характер браков должен был привести к максимальному сходству, а не к различиям южных популяций. Поэтому много более вероятно, что большие генетические различия связаны не с реальной структурой популяции, а с характером формирования выборок. Если часть обследованных не являлась в строгом смысле представителями коренного населения, то в выборке могли появиться гаплогруппы, необычные для южных русских популяций, и эти «залётные» гаплогруппы могли оказаться самыми разными в каждой из обследованных групп. Рассмотрим, как частоты гаплогрупп в этих трёх популяциях отличаются от среднерусских частот (средних частот в популяциях, вошедших в основной кластер нарис.6.2.1.). КРАСНОДАРСКАЯ ВЫБОРКА отличается повышенной частотой гаплогруппы J (18% в Краснодарской популяции при среднерусской частоте 9%) и сниженной частотой гаплогруппы U (10% в Краснодарской популяции при среднерусской частоте 19%). При этом частота гаплогруппы J у коренных народов Северного Кавказа (в среднем 8%) вовсе не повышена, а близка к среднерусской. А частота гаплогруппы U у народов Северного Кавказа (25%) отнюдь не ниже, а даже выше среднерусской (19%). То есть отличия Краснодарской выборки от остальных русских популяций нельзя объяснить смешением с народами Кавказа. Остается предполагать, что в эту выборку попали представители каких-то третьих групп, или же своеобразных, нетипичных популяций. СТАВРОПОЛЬСКАЯ ВЫБОРКА отличается от основного кластера русских популяций сниженной частотой гаплогруппы Н (35% вместо 42%) и повышенной частотой гаплогруппы X (8% вместо 1%). В данном случае смешение с местными северокавказскими популяциями отчасти могло сформировать такие частоты (средняя частота гаплогруппы Н на Северном Кавказе - 22%). Хотя главную особенность Ставропольской выборки - совершенно необычную частоту X - трудно объяснить смешением, поскольку и на Северном Кавказе такая высокая частота X встречается лишь в немногих популяциях, а её средняя частота на Кавказе - 5%. Чтобы объяснить высокую частоту X в русском населении Ставрополья, придётся предположить, что этот генофонд большей частью сформирован материнскими линиями не из Центральной или Южной России, а автохтонного кавказского населения, причём каких-то специфических групп. Очевидно, что такое предположение неправдоподобно. Вернее всего, что сам генофонд Ставропольского края вполне сходен со среднерусским, но конкретная рассматриваемая выборка плохо отражает генофонд Ставрополья. Впрочем, это легко можно будет проверить, когда появятся данные ещё по одной-двум русским популяциям из этого региона. ВЫБОРКА КУБАНСКИХ КАЗАКОВ. Что же касается третьей популяции из Предкавказья - кубанских казаков (старожильческого русского населения Адыгеи), то эта популяция оказывается вполне «среднерусской». Ни по одной из гаплогрупп отклонения от среднерусских частот не превосходят 5%. Поэтому и на графике она разместилась вблизи основного кластера русских популяций (рис. 6.2.1.). А ведь именно в этой из трёх южных популяций и можно было ожидать максимального своеобразия - и вследствие большей давности их отделения от основной части русского генофонда, и вследствие их проживания вперемешку с коренным населением Северного Кавказа. В следующей главе мы покажем, сколь своеобразна эта популяция по данным о фамилиях. Поэтому близость кубанских казаков по мтДНК к среднерусским частотам лучше всего показывает истинное положение и других южных популяций в генетическом пространстве. Небезынтересно, что как раз эта выборка кубанских казаков (наши данные) была собрана Э. А. Почешховой наиболее тщательно - собирались сведения о местах рождения родителей и бабушек-дедушек обследованных. Если рассматривать предков только по материнской линии, то выборка главным образом (60%) состоит из уроженцев Майкопского района республики Адыгея, на 36% - из уроженцев других районов Адыгеи и Краснодарского края и лишь оставшиеся 4% бабушек со стороны матери происходят из Ростовской, Воронежской областей и Ставропольского края. Мы видим, что выборка, которая более чем на~95% и на глубину трёх поколений представляет русское казачье население Краснодарского края, близка к общерусскому кластеру! А выборки, собранные менее тщательно в том же Краснодарском и в Ставропольском краях, располагаются весьма далеко от основного массива русских популяций. Итак, необычные частоты гаплогрупп в Краснодарской и Ставропольской выборках задаются случайностями выборки, а не своеобычностью самой популяции. Третья - корректно сформированная - южная выборка оказалась близка к большинству русских популяций. ВСЕГДА ЛИ ПОПУЛЯЦИИ РАЗЛЕТАЮТСЯ? На графике (рис. 6.2.1.) есть и другие примеры, когда из одного региона изучено несколько популяций. Например, две выборки из Белгородской области (Белгород 1 и Белгород 2), независимо собранные разными исследователями (см. табл. 6.2.1) оказались генетически близки. Обе северно-русские популяции (Пинега и Каргополь) отличаются от основного кластера русских популяций повышенными частотами гаплогрупп Н, К, и U5b, сниженными частотами гаплогруппы U5a, и, по всей видимости, V. Важно, что эти обе северно-русские популяции на графике близки друг к другу. В противном случае их особенности можно было бы толковать как результат хаотического действия дрейфа генов. Однако трудно предположить, что дрейф, действуя на две популяции (разнесённые на 400 км!) сместил частоты одних и тех же гаплогрупп в одну и ту же сторону. Можно надеяться, мы имеем дело с реальным генетическим портретом населения Русского Севера. ГЕТЕРОГЕННОСТЬ РУССКОГО ГЕНОФОНДА Рассматривая «митохондриальный генофонд» русских популяций, надо количественно ответить на вопрос: какова гетерогенность русского генофонда? Велики или малы генетические различия между русскими популяциями по данным о мтДНК? Средние генетические расстояния между семью русскими популяциями составили d=0.033. Это и есть оценка гетерогенности русского генофонда по мтДНК: величина различий русских популяций между собой. МЕТОДИКА РАСЧЕТА. Для оценки различий использовались генетические расстояния Нея, рассчитанные в программе DJ, написанной Ю. А. Серегиным (программа представлена на нашем сайте www.genofond.ru). Исходные данные представлены частотами 21 гаплогруппы: Н; HV; HVpre; I; J; К; N1; R; T; U1; U2; UЗ; U4; U5a; U5b; U7; U8; V включая Vpre; W; X; а также суммой восточно-евразийских гаплогрупп А, В, С, D, G, Y. Мы рассчитали генетические расстояния между каждой парой русских популяций и величину средних генетических расстояний. Генетический смысл средних расстояний тот же, что и показателя межпопуляционной изменчивости GST - оба показателя оценивают размах различий между популяциями [Bodmer, Cavalli-Sforza, 1976]. ВЫБОРКИ РЕШАЮТ ВСЕ! (ИЛИ МНОГОЕ...) Отметим, что если наша оценка дифференциации русских популяций по мтДНК составила d=0.033, то по данным Б. А. Малярчука она порядок ниже: d=0.002. По всей видимости, столь серьёзные расхождения возникли оттого, что в работе Б. А. Малярчука изучены популяции лишь из южной части русского ареала (Ставропольского и Краснодарского краев. Белгородской, Орловской, Саратовской и Нижегородской областей), тогда как в нашем исследовании юг и север, запад и восток представлены достаточно равномерно. По-видимому, межпопуляционные различия между южными популяциями действительно малы, что и было обнаружено Б.А. Малярчуком. Но, как мы видели (рис. 6.2.1), генетические различия между русским югом и русским севером по мтДНК очень велики и они определяют большую изменчивость русского генофонда в целом. В работе [Малярчук, Деренко, 2004] приведён показатель дифференциации FST, который по частотам 29 гаплогрупп составил FST=-0.0006, а по частотам гаплотипов ГВС1 FST=-0.0016. Авторы сделали вывод: «. ..Полученные результаты свидетельствуют об отсутствии генетической подразделённости по митохондриальным линиям между изученными популяциями русского населения...». Тем удивительнее полученная нами высокая оценка гетерогенности. Ведь FST с небольшой долей огрубления можно приравнять к использованному нами показателю - средним генетическим расстояниям (выше мы говорили о сходстве расстояний с третьим показателем - GST очень близким к FST). Этот пример показывает, насколько важен охват самых различных частей ареала. Иначе легко сделать выводы, верные лишь для фрагмента ареала. Отметим, что в одной из недавних работ Б. А. Малярчук (с учетом новых полученных им данных по более широкому кругу популяций) оценил дифференциацию русского генофонда как значительную [Malyarchuk et al., 2004]. РУССКИЙ ГЕНОФОНД НА ФОНЕ НАРОДОВ ЕВРОПЫ Имеющиеся данные показывают, что гетерогенность других европейских народов по мтДНК значительно меньше. Гетерогенность мтДНК русского генофонда по своему размаху соответствует скорее гетерогенности не отдельных европейских народов, а целых регионов Европы. Например, рассчитанные нами различия по мтДНК между всеми локальными популяциями в пределах_ Балканского региона составили ту же величину (d=0.033), что и различия между всеми русскими популяциями, а различия между локальными популяциями всей Западной Европы (от Испании до Скандинавии) лишь ненамного выше (d=0.041). ИТОГИ Итак, мы рассмотрели расположение в генетическом пространстве мтДНК всех двенадцати русских популяций, изученных к настоящему времени. Большинство популяций, независимо от своего географического положения, формируют единый кластер в генетическом пространстве. Особое положение занимают северные русские популяции, что, вероятно, отражает реальные особенности генофонда Русского Севера. Что же касается двух южных «разлетевшихся» популяций, то их поведение, по-видимому, зависит лишь от случайностей формирования этих выборок. Средние генетические расстояния между всеми русскими популяциями (d=0.033) свидетельствуют о высокой гетерогенности русского генофонда на общеевропейском фоне. Таковы основные итоги обобщенного анализа мтДНК на уровне гаплогрупп. Остаток «митохон- дриального» раздела данной главы мы посвятим анализу в другом масштабе - гаплотипов. §4. Сказка об изолах и миграхЖИЛИ-БЫЛИ ИЗОЛЫ И МИГРЫ... В конце палеолита племя изолов укрылось в кольце непроходимых гор. Прошли тысячелетия, но эта популяция сохранилась в полной изоляции. На их высокогорном плато нет природных барьеров и велика подвижность населения - они охотники и скотоводы. Эти особенности образа жизни не позволили изолам разделиться на устойчивые подгруппы. В результате все изолы имеют примерно равные шансы вступить в брак друг с другом (рис. 6.2.2). А рядом с изолами - по ту сторону гор - в исторически недавнее время поселились мигры (рис. 6.2.2). Численность мигров и изолов одинакова. Но народ мигров составился из четырех разных племен, подобранных ими на долгом пути миграции. Так мигры и расселились - каждое племя в своей долине. Основная доля браков заключается в пределах племени. Но по большим праздникам устраиваются всеобщие ярмарки, где порой заключаются и межплеменные браки, и браки с другими народами. Итак, народ мигров - подразделённая популяция, образовавшаяся совсем недавно. Изолы - бесструктурная (панмиксная) и очень древняя популяция. Изолы произошли от палеолитической популяции, имевшей лишь несколько древних гаплотипов мтДНК. С течением времени благодаря мутациям возникали все новые гаплотипы, нарастали молодые побеги родословного древа гаплотипов. Росло генетическое разнообразие внутри популяции. Так действовали мутации. Но второй, более мощный фактор - дрейф генов - это разнообразие уменьшал, случайно оставляя лишь малую часть гаплотипов, не считаясь с тем, новорожденные эти гаплотипы или очень древние. Но эти редкие случайно «выжившие» гаплотипы встречаются с высокой частотой, и вновь частота их может не зависеть от того, новорожденные эти гаплотипы или очень древние. Ведь дрейф работает «вслепую», безответственно и неотразимо, не считаясь с родословными. Мигры же, собрав по пути множество гаплотипов, принесли их на свою новую родину. С тех пор прошло всего несколько поколений, и новые гаплотипы ещё не возникли. Зато племена мигров по-прежнему открыты внешним миграциям, пополняющим разнообразие гаплотипов. РАЗНЫЕ ИТОГИ ДРЕЙФА У ИЗОЛОВ И МИГРОВ Ни изолы, ни мигры не могут избежать действия дрейфа генов, истребляющего, один за другим, гаплотипы из их популяций. То, что дрейф именно так поступает с редкими генами, популяционная генетика открыла очень рано, и это стало хрестоматийным примером. Причём тогда генетики работали с аутосомными генами, а не с мтДНК. Это важное различие: ведь мтДНК, во-первых, гаплоидна (не две, как для аутосомных генов, а только одна копия на геном человека), а во-вторых, передаётся только от половины родителей (от матерей). Перемножая эти величины, получаем, что число митохондриальных ДНК, переданных следующему поколению, в четыре раза меньше, чем для любого аутосомного гена. Поэтому дрейф действует на мтДНК примерно в четыре раза сильнее, эффективнее пропалывая гаплотипы. Но дрейф по-разному действует на изолов и на мигров. В панмиксной популяции изолов действие дрейфа непоправимо: исчезнувший гаплотип никогда не вернется. А в подразделённой популяции мигров гаплотип, исчезнувший в одном из племен, может снова прийти из другого племени. Вероятность того, что этот гаплотип независимо исчезнет во всех четырех племенах, мала. Случайное уменьшение частоты в одной из частей подразделённой популяции может компенсироваться её случайным увеличением в другой, и в результате средняя частота у мигров более устойчива. Это явление стабильности подразделённой популяции описано во многих работах и подробно рассмотрено в Приложении. МУХИ. Наиболее прямые доказательства устойчивости подразделённых популяций к дрейфу генов получены в известных работах Юрия Петровича Алтухова и Елены Юрьевны Победоносцевой, которые провели прямой эксперимент на дрозофиле. Мухи были расселены по отсекам популяционного ящика и обменивались генами точно так, как мы предположили для мигров - равная численность в отсеках, равновероятный обмен между отсеками, но основная часть скрещиваний происходит в отсеке. Исходная частота гена была 0.5 и оставалась практически неизменной: в течение 80 поколений варьировала от 0.6 до 0.4. (см. Приложение). Другой ящик тех же мух, с той же обшей численностью и в тех же условиях, но с полной свободой скрещивания, - моделирует панмикснуто популяцию изолов. В ней - за то же время - частота гена упала от 0.6 до 0.1 и этот ген легко может быть потерян. Итак, подразделённая популяция устойчива и хранит генетическую память. А панмиксная - нет. Мы убедились, что они эволюционируют с разной скоростью. ЛЮДИ. Конечно, история изолов и мигров сложнее этого эксперимента. В ней случались и засухи, и эпидемии, и войны. Порою численность увеличивалась, а потом резко падала. Это приводило к эффекту «бутылочного горлышка». Для генетической памяти мигров угроза невелика - подразделённая популяция обладает особым свойством репарации. Пусть даже одно из племен полностью исчезнет, средние частоты народа в целом изменятся незначительно. А на ареал исчезнувшего племени вернутся их отдалённые потомки - дети от межплеменных браков (земля предков всегда манит потомков). И тогда генофонд исчезнувшего племени будет отчасти восстановлен. Однако для генетической памяти панмиксной популяции изолов эффект «бутылочного горлышка» очень опасен, даже если численность потом восстановилась. При колебаниях численности дрейф генов действует ещё разрушительнее, так как средний по всем долгим поколениям эффективный размер популяции Ne близок к «диаметру бутылочного горлышка», а дрейф генов целиком и полностью зависит от эффективного размера популяции (см. Приложение). ИЗУЧАЕМ ИЗОЛОВ И МИГРОВ... И ДЕЛАЕМ НЕВЕРНЫЕ ВЫВОДЫ! Какие же результаты получим мы, изучив современных изолов? Дрейф генов приведёт к потере большинства гаплотипов - как древних, так и новых мутаций. То, какие именно гаплотипы мтДНК или Y хромосомы будут утеряны, - зависит от чистой случайности. Следовательно, от неё зависят и наши выводы. Если уцелеет группа гаплотипов, несущих общую позднюю мутацию, - мы сделаем вывод о чрезвычайной молодости изолов. Если уцелеют две разные группы гаплотипов - мы сделаем вывод о смешении двух разных народов. И это когда перед нами самый древний в мире народ - прямой потомок палеолита, который за 20 тысяч лет своей истории не принял ни одного чужеродного гена! А что же мигры? Популяция мигров, благодаря подразделённости, сохраняет феерическое разнообразие гаплотипов. Это именно та картина, которую представляла бы древнейшая популяция изолов, если бы на них не действовал дрейф. То есть мигры предстанут перед исследователем блестящей имитацией древней популяции. Итог изучения мтДНК может быть таким: мигры - древнейшее коренное население края, а изолы - недавняя миграция неизвестно откуда. То есть прямая противоположность реальной истории. В ЧЁМ ЖЕ ОШИБКА? И эту злую шутку сыграло с нами игнорирование структуры популяций. Несомненно, в генофонде и изолов, и мигров записана их история. Но эти записи, переписываемые в каждом поколении, могут искажаться. У популяции есть генетическая память. Но подразделённые мигры имитируют чужую память, а панмиксные изолы свою память утратили. В древних китайских хрониках о варварах говорилось, что они не помнят прошлого, не думают о будущем. И применительно к генофонду, его «цивилизованность» (твёрдая генетическая память) или «варварство» (её отсутствие) оказываются тесно связанными с подразделённостью популяции, её структурой. СКАЗОЧНЫЕ УРОКИ Эта сказка показывает, что для интерпретации результатов изучения мтДНК недостаточно представления о том, что разнообразие гаплотипов в популяции, благодаря мутациям, увеличивается с ходом времени. Для правильной интерпретации необходимо также знать: какова структура популяции? является ли популяция коренной? а если смешанной и пришлой, то где и как она формировалась? могла ли сохраниться генетическая память? 
§5. Прокрустово равенство: сколько «русских» гаплотипов у соседей по Европе?Миновав Элевсин и приближаясь уже к долине реки Кефиса в Аттике, Тесей пришёл к разбойнику Дамасту, которого называли обыкновенно Прокрустом... Прокруст, легендарный разбойник, имел ложе, в которое укладывал свои жертвы: если их рост был меньше длины ложа, он их растягивал, если больше - отрубал лишнее. Мифы и легенды Древней Греции, Греческие крылатые выражения Дальше им пришлось миновать пролив, по одну сторону которого подстерегала корабли ужасная Харибда, а по другую - чудовищная Сцилла. Харибда выглядела подобно водовороту, втягивавшему в пасть воду вместе с кораблями. Сцилла же была некогда прекрасной девой, и злая Кирка, завидуя ей, превратила её в страшилище с шестью собачьими головами и двенадцатью ногами. Одиссей направил корабль мимо последней, решив пожертвовать шестью спутниками, так как понимал, что Харибда проглотит корабль целиком. Мифы Древней Греции ЧТО ТАКОЕ ГАПЛОТИПЫ? Мы понимаем под гаплотипом вариант мтДНК, который можно отличить от других таких же вариантов - неважно, различаются они по ГВС1 или же по ПДРФ маркёрам кодирующей части. Среди изученных нами 650 индивидуумов из исторического ареала русского народа выявлено 283 различных гаплотипа. Если добавить к ним все опубликованные и архивные данные по русским, то общая выборка составит 1584 образцов, а число гаплотипов в ней - 628. Это одна из самых больших выборок по мтДНК среди всех народов мира. ВОЗМОЖНОСТИ ГАПЛОТИПОВ Огромное разнообразие гаплотипов придаёт сравнению популяций потенциально большую разрешающую способность. Ведь мы можем обнаружить как гаплотипы, общие между двумя популяциями, так и те гаплотипы, которые присутствуют только в одной из них. Можем рассмотреть, присутствует ли данный гаплотип в смежных популяциях и в каких именно. И проведем мы такой анализ не по десятку признаков (как в случае аутосомных генов или гаплогрупп мтДНК), а по сотням разных признаков (гаплотипов мтДНК). Например, мы рассмотрим гаплотипы, встреченные у русских и у соседних народов. И те популяции, с которыми у русских выявится больше общих гаплотипов, сочтём наиболее родственными. НЕНАСЫТНАЯ ХАРИБДА: А ВСЕ ЛИ ГАПЛОТИПЫ НАЙДЕНЫ? Несомненно, в русском населении существует некое конечное число гаплотипов. И при достаточно большой выборке они могут быть описаны почти все. Вопрос лишь в том, насколько велика должна быть такая выборка? Этот вопрос вызван отнюдь не праздным любопытством. Для описания митохондриального генофонда популяции хотелось бы знать, какие именно гаплотипы там существуют, иметь их перечень. Это особенно важно для сравнения популяций. Если мы не знаем полного перечня гаплотипов, существующих в двух популяциях, то любые различия могут быть вызваны тем, что в одну из выборок эти гаплотипы просто не попали, хотя в популяции они присутствуют! Иными словами, при сравнении популяций по редким признакам (например, гаплотипам мтДНК) особенно важно знать, в какой степени наша выборка описывает реальную популяцию. Этот вопрос критичен и для практических целей судебно-медицинского использования мтДНК: «если данный гаплотип в популяции не обнаружен - то какова вероятность, что он там все-таки есть?» ЧЕМ БОЛЬШЕ ВЫБОРКА. ТЕМ БОЛЬШЕ ГАПЛОТИПОВ Чтобы ответить на этот вопрос, мы изучили скорость накопления гаплотипов при увеличении объёма выборки. Как можно видеть на графике (рис. 6.2.3.), число гаплотипов в русских популяциях возрастает с увеличением объёма выборки линейно. И мы должны констатировать, что признаков насыщения не обнаружено. Насыщение должно было бы проявиться в постепенном выходе кривой на плато или, по крайней мере, в замедлении роста числа гаплотипов при увеличении выборки. Такая «ненасытность» приводит к весьма важному выводу: реальное число разных линий мтДНК в больших и географически подразделённых популяциях очень велико. Даже при выборках в тысячу и более образцов, по-прежнему в популяции остается множество невыявленных гаплотипов. Эту особенность мтДНК всегда необходимо учитывать, сравнивая популяции по спектру гаплотипов. Ведь если какой-либо гаплотип встречен у русских, но не обнаружен в иной популяции, это отнюдь не означает реального различия этих популяций. В действительности гаплотип может быть в обеих популяциях, но выявим мы его только при увеличении выборки. 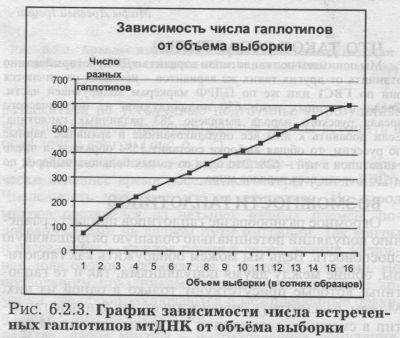
КАК ПОСТРОЕН ЭТОТ ГРАФИК. Из общего массива в 1600 образцов из русских популяций случайным образом выбиралось по 100 образцов, и подсчитывалось число гаплотипов. На следующем шаге добавлялось ещё 100 образцов, случайно выбранных из оставшихся 1500, и подсчитывалось число гаплотипов в этой увеличившейся выборке. Всего было сделано 16 таких шагов - пока реальные доступные данные (1600 образцов) не были исчерпаны. Грубая аппроксимация показывает, что соотношение между общим объёмом выборки и числом различных линий (гаплотипов) составляет примерно 7:3. Иными словами, число гаплотипов составляет около 40% от числа исследованных образцов русского народа. Мы проверили это соотношение по данным о другом хорошо изученном народе - немцам, где исследовано лишь немногим меньше образцов - 1396. Число разных гаплотипов в этой выборке немцев составило 553, то есть соотношение осталось тем же - 40%. Конечно, это соотношение должно несколько меняться в зависимости от разрешающей способности филогенетического анализа, то есть нашей способности отличать друг от друга «похожие» гаплотипы. Это соотношение должно зависеть и от структуры популяции, от соотношения миграций и дрейфа генов. Например, для саамов (лопарей), которые служат классическим примером мощного дрейфа, доля разных гаплотипов составляет, по нашим расчётам, только 11% от объёма выборки (63 гаплотипа в общей выборке 559 человек, информация Банка данных World Mitochondrial, см. Приложение раздел 5.). КАК СРАВНИТЬ ПОПУЛЯЦИИ? Сравнивать частоты гаплотипов, по всей видимости, нецелесообразно - слишком низка частота практически любого гаплотипа. И двукратные различия в частотах (в какой-то популяции один, а где-то встретилось два человека с таким гаплотипом) будут вызваны лишь случайностями выборки. Лучше сравнивать на качественном уровне: не важно, у скольких индивидов встречен данный гаплотип - важно, что он обнаружен в популяции. Тогда зависимость от объёма выборки будет меньше. РАСЧЁТ ИНДЕКСА СХОДСТВА. Для каждой пары сравниваемых популяций мы подсчитывали число общих филогенетических линий (гаплотипов) мтДНК. Индекс сходства оценили как соотношение (долю общих линий): в числителе - количество линий мтДНК, общих с русскими популяциями; в знаменателе - суммарное число линий мтДНК, обнаруженных в сравниваемой популяции. Такой способ расчёта, на первый взгляд, решает проблему «нерепрезентативности» любых наших выборок (в том смысле, что в них представлены далеко не все гаплотипы, реально присутствующие в популяции). Решает за счет того, что мы подсчитываем не число, а долю гаплотипов, общих между двумя популяциями. Ведь все гаплотипы имеют одинаковую вероятность «потеряться» (присутствовать в популяции, но отсутствовать в выборке). И если для изучаемой популяции «А» общие гаплотипы обнаруживаются преимущественно с популяцией «В», и реже с популяцией «С», то это означает, что таково реальное соотношение митохондриальных генофондов этих популяций. При этом мы предполагаем, что при увеличении объёма выборок число общих гаплотипов должно увеличиться в каждой сравниваемой паре популяций, но само соотношение останется примерно тем же. КОВАРНАЯ СЦИЛЛА: В ЧЁМ ОШИБАЕТСЯ ИНДЕКС СХОДСТВА? К сожалению, проверка показывает, что это не1 вполне так. На рис. 6.2.4. приведена зависимость доли общих гаплотипов от объёма выборки (нижняя линия). Ясно видно, что такая зависимость имеется: чем меньше объём выборки, тем больше индекс сходства, тем больше доля общих гаплотипов. ВЕРОЯТНОЕ ОБЪЯСНЕНИЕ. Можно предположить, что это вызвано наличием в популяциях сравнительно частых гаплотипов. Например, для населения Европы такими гаплотипами окажутся: в гаплогруппе Н гаплотип CRS и гаплотип с заменами «304311»; в гаплогруппе J - гаплотип с заменами «069-126». Эти гаплотипы очень часты в Европе в целом, и поэтому встречены почти в каждой изученной европейской популяции. Число таких гаплотипов невелико. Но именно за счет своей повсеместности они всегда окажутся общими между любыми популяциями. Тем самым они являются как бы постоянной «прибавкой» к числу общих гаплотипов. И чем меньше объём выборки, тем больше сказывается это постоянное прибавочное число, тем больше оно завышает долю общих гаплотипов и повышает индекс сходства популяций. Примечание. Для десяти европейских популяций подсчитаны три показателя: общее число гаплотипов в них; число гаплотипов, общих с русскими популяциями; доля гаплотипов, общих с русскими популяциями. Все 10 рассматриваемых популяций характеризуются разными объёмами выборок. График показывает, как три перечисленных параметра (отложенных по оси ординат) зависят от объёма выборки (ось абсцисс). Общее число гаплотипов при увеличении объёма выборки возрастает. Число гаплотипов, общих с русскими, также возрастает. Доля гаплотипов, общих с русскими, снижается. Это снижение хорошо видно на аппроксимирующем графике (жирная пунктирная линия). Для совместимости на одном графике число общих гаплотипов приводится умноженное на 10, а доля - умноженная на 1000. Справедливо ли такое объяснение или есть другие причины зависимости сходства популяций от объёма выборки, в любом случае из-за этой зависимости любые выводы повисают в воздухе. Достаточно увеличить выборку, и обнаруженное ранее сходство популяций исчезнет! Наименее похожими на русских окажется население тех стран, где генетические исследования проводятся более интенсивно, а наиболее сходными будут просто слабо изученные народы! Необходим способ расчёта, корректирующий эту зависимость. 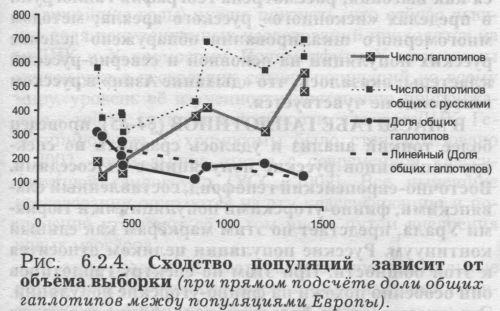
ПРОКРУСТ СПЕШИТ НА ПОМОЩЬ Нами использован, пожалуй, самый простой, прямой, а потому надёжный способ такой коррекции - Прокрустов. Все сравнения проведены на выборках одного и того же объёма. Чтобы выровнять объёмы выборок, из каждой случайным образом было отобрано одно и то же число образцов, и дальнейшие сравнения проводились именно с этими новыми выборками одинакового объёма. Естественно, при этом происходила потеря части данных - все выборки приходилось выравнивать по самой маленькой из них (по размеру прокрустова ложа). Тем не менее, только такой прокрустов подход («отрубать лишнее») позволяет нам быть уверенными, что полученные результаты не зависят от того, что какая-то популяция изучена по большей выборке, а отражают что-то более объективное, можно надеяться - реальное сходство популяций. СРАВНЕНИЕ РЕГИОНОВ. А НЕ ЛОКАЛЬНЫХ ПОПУЛЯЦИЙ Зная, какие гаплотипы встречены в каждой из русских популяций, можно было бы сравнить разные русские популяции и определить, какие из них наиболее сходны друг с другом. Похожий анализ мы проводили по данным о частотах гаплогрупп (§3). Но осторожность не позволяет нам сделать это по данным о гаплотипах: выборки по каждой популяции составляют 100-150 человек, что слишком мало для определения спектра гаплотипов. Поэтому мы не можем провести анализ внутри русского генофонда, сравнив между собой разные русские популяции. Но можем объединить данные по всем русским популяциям, получив многосотенную выборку. И сравнить такой общерусский спектр с аналогичными спектрами соседних групп населения (табл. 6.2.4.). НА КОГО МЫ ПОХОЖИ? 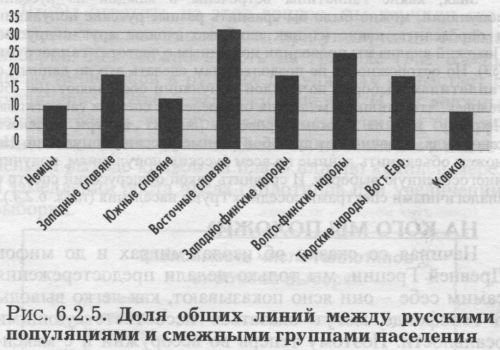
Начиная со сказки об изолах-миграх и до мифов Древней Греции, мы только делали предостережения самим себе - они ясно показывают, как легко выводы о генофонде могут оказаться несоответствующими реальности. Поэтому теперь во всеоружии и с максимальной осторожностью сравним спектр гаплотипов в русских популяциях и у их соседей по Европе. Диаграмма (рис. 6.2.5.) показывает, что по степени сходства с русским генофондом его соседи располагаются в следующем порядке: восточные славяне, волго-финские народы, западно-финские народы, западные славяне, тюркские народы Урала, южные славяне, германоязычные народы, народы Кавказа. То, что по спектру гаплотипов русские оказались похожи на прочих восточных славян, не удивительно. Однако западные и в особенности южные славяне заметно отличаются от русских. У финноязычных народов (как западных, так и волжских) несколько больше общих гаплотипов с русскими, чем у славянских народов! Это можно трактовать в терминах финно-угорского субстрата в русском населении. Тюркоязычные народы и население Кавказа, несмотря на тесное соседство с современной зоной расселения русских, имеют с ними меньше общих гаплотипов мтДНК. Эти результаты показывают общность митохондриального генофонда Восточной Европы. Во всех языковых группах этого региона (восточные славяне, финно-угры, тюрки Уральского региона) мы видим общий массив гаплотипов мтДНК. Все эти группы народов по спектру гаплотипов похожи на русские популяции и, соответственно, должны быть и похожи друг на друга. На Кавказе и в Западной Европе спектр гаплотипов уже несколько отличается. Западные славяне, судя по рис. 6.2.5., присоединяются к восточноевропейскому кластеру, а южные славяне отличаются от него так же, как Кавказ и Западная Европа. Сравнительно низкая доля общих линий между русскими и южными славянами требует объяснения. Можно вспомнить, что в ходе славянской экспансии как восточные, так и южные славяне расселились на территориях со значительно отличающимся субстратным населением: финноязычным на востоке и иллирийским на юге. Смешение с этими различными субстратами могло привести к заметным различиям между двумя славянскими группами по мтДНК. Нельзя также исключать, что предковые группы южных и восточных славян были исходно генетически различными. ОСНОВНЫЕ ЧЕРТЫ «МИТОХОНДРИАЛЬНОГО ПОРТРЕТА» В разделе 6.2. мы рассмотрели изменчивость «митохондриального генофонда» в двух масштабах. В МАСШТАБЕ ГАПЛОГРУПП (§2-§3): гетерогенность русского генофонда по мтДНК оценивается как высокая; рассмотрена география гаплогрупп в пределах «исконного» русского ареала; методом многомерного шкалирования обнаружено деление русских популяций на основной и северно-русский кластеры; оказалось, что «дыхание Азии» в русском генофонде не чувствуется. В МАСШТАБЕ ГАПЛОТИПОВ (§4-§5): проведен более тонкий анализ и удалось сравнить по спектру гаплотипов русские популяции с их соседями. Восточно-европейский генофонд, составленный славянскими, финно-угорскими популяциями и тюрками Урала, предстает по этим маркёрам как единый континуум. Русские популяции целиком относятся к этой общности. При этом по спектру гаплотипов они особенно похожи на финно-угорские популяции. Это указывает на большую роль финно-угорского субстрата в формировании митохондриального генофонда русских популяций. «МИТОХОНДРИАЛЬНЫЙ ПОРТРЕТ» рисует картину русского генофонда, близкую к тем портретам, которые были получены и по данным антропологии, и по классическим генетическим маркёрам. Данные по мтДНК также показывают чётко выраженные различия между южными и северными русскими группами. Размах этих различий заметен даже в общеевропейском масштабе изменчивости, но нигде не выходит за его рамки. Прослеживаются особенности генофонда, отражающие влияние смежных групп, в особенности финно-угорских популяций. Подробнее итоги изучения мтДНК описаны в разделе 6.4. 
2Пока эта глава готовилась к печати, в конце 2004 года вышла ешё одна статья Б. А. Малярчука с новыми данными по русским популяциям [Malyarchuk et al., 2004]. Эти популяции, к сожалению, уже не могли быть включены в наш анализ, но чтобы дать читателю полный перечень изученных русских популяций, мы приводим их в таблице 6.2.2. 3Аутосомные гены «диплоидные», т. е. присутствуют в двойном наборе. А у «гаплоидных» однородительских маркёров лишь один набор генов - либо только от матери (мтДНК), либо только от отца (Y хромосома). Поэтому выборка людей для изучения однородительских маркёров и должна быть в два раза больше. Ведь изучив 75 человек по аутосомным маркёрам, мы получаем выборку 150 генов. А по однородительским для выборки 150 генов мы должны изучить 150 человек. 4В популяциях имеется множество гаплотипов, поэтому их частота невелика. И надо увеличивать численность выборки из популяции - тогда появляется шанс, что они в неё попадут. Если частота исчисляется сотыми долями, то и выборка должна быть не меньше 100 человек. |
загрузка...