6.1. Аутосомные ДНК маркёры
|
СПИД протекторная мутация - Её большое значение - Её любопытная история - Элегантность её географии - Распространение в русском ареале - Западнорусская аномалия
Выше мы говорили о том, насколько важно для изучения пространственной структуры русского генофонда охватить популяции из различных частей его ареала. К сожалению, география изученных популяций надёжна лишь для немногих аутосомных ДНК маркёров. Пожалуй, лучше всего в русском ареале изучено распределение мутации в гене CCR5. Поэтому рассмотрим географию именно этого популярного ДНК маркёра. В последние десять лет эта мутация - делеция 32 нуклеотидов в гене рецептора хемокинов (CCR5del32) - привлекает большое внимание исследователей всего мира. Причина очевидна: гомозиготы по мутантному гену CCR5del32 почти полностью устойчивы к инфицированию ВИЧ-1. Имеются данные для 22 русских популяций (табл. 6.1.1). Правда, для трёх из них географическая привязка или отсутствует [Stephens et al., 1998], или проблематична [Libert et al., 1998], или характеризует население за пределами «исконного» ареала (русские Новосибирска [Yudin et al., 1998]). Но остальные 19 популяций позволяют картографировать распространение мутации CCR5del32 достаточно подробно. МЕХАНИЗМ УСТОЙЧИВОСТИ К ВИЧ-1 Молекулярные механизмы такой устойчивости хорошо изучены [Dragic et al, 1996; Dean et al, 1996; Samson et al, 1996]. Вирус использует хемокиновый рецептор для проникновения в клетку. Мутация приводит к отсутствию рецептора на поверхности клетки. а это препятствует проникновению вируса. В то время как гомозиготные носители делеции обладают почти полной устойчивостью, у гетерозиготных пациентов СПИД все-таки развивается, но медленнее, на два-три года позже, чем обычно [Zimmerman et al, 1997; Galvani, Novembre, 2005]. Кроме del32, в гене CCR5 и прочих генов семейства хемокинов (CCR2, SDF1) обнаружены и другие мутации. Некоторые из них тоже снижают темпы инфицирования и относятся к СПИД протекторным генам, но их влияние слабее CCR5del32 [O'Brien, Moore, 2000]. ГЕОГРАФИЯ И ВОЗРАСТ ДЕЛЕЦИИ Известно, что мутация CCR5del32 встречается главным образом у европейцев (в среднем 10%). Частота её в Европе снижается с севера на юг. За пределами Европы мутация встречается с низкими частотами в смежных регионах (Северная Африка, Ближний Восток, Средняя Азия), но отсутствует в более отдалённых от Европы регионах: в Африке южнее Сахары, в Восточной и Юго-Восточной Азии, в коренном населении Америки [Martinson at al, 1997, 2000]. Такой простой тренд необычен для распространения генов в популяциях человека. И он стал особенно интригующим, когда появились данные по возрасту мутации. Основываясь на изменчивости микросателлитных маркёров, сцепленных с геном CCR5, Либерт с коллегами [Libert at al, 1998] показали, что мутация возникла примерно 2000 лет назад. Данные другой группы [Stephens et al, 1998] оценили возраст мутации всего в 700 лет. Впрочем, 95% доверительные интервалы в обоих случаях покрывают несколько тысячелетий. Тем не менее, нет сомнений, что мутация возникла лишь однажды и возникла в Европе. Следовательно, частота мутации в населении Европы возросла от нуля до 10% за сравнительно короткое время. Согласно законам популяционной генетики, такое резкое изменение частоты в столь большой популяции не может быть вызвано случайным дрейфом генов [см. обзор Galvani, Novembre, 2005]. Остается предполагать действие естественного отбора в пользу этой мутации. Это сразу возникающее впечатление - действия отбора в пользу мутации - было подтверждено и математическим моделированием [Stephens et al, 1998]. Эпидемия СПИДа началась слишком недавно, чтобы изменить частоты генов. Следовательно, нужно найти иные факторы естественного отбора, действовавшие в историческое время. Опираясь на тот факт, что хемокиновые рецепторы взаимодействуют, кроме ВИЧ-1, и с другими патогенами, было выдвинуто предположение, что мутация может определять устойчивость сразу к нескольким инфекционным заболеваниям. ГИПОТЕЗЫ «ГИПОТЕЗА ЧУМЫ» [Stephens et al., 1998] постулирует, что частота мутации возросла в результате эпидемий чумы. Гипотеза основана на том, что в средневековой Европе от чумы погибала заметная часть населения. Это должно было привести к заметному действию естественного отбора в пользу любого фактора, определяющего устойчивость к болезни. Эта гипотеза, хотя и получила широкое распространение, никогда не имела прямых доказательств. Более того, недавно было показано, что мутации в гене CCR5 не влияют на протекание чумы у мышей [Mecsas et al., 2004]. «ГИПОТЕЗА ОСПЫ». Но, возможно, что инфекционным заболеванием, приведшим к распространению мутации, была оспа [Galvani, Slatkin, 2003]. Эпидемии оспы, хотя были не столь драматичны, как чума, но поражали европейцев непрерывно. Причём жертвами обычно были дети, что могло ускорить изменение частоты аллеля в поколениях. Опять-таки, и эта гипотеза основывается не на клинических или молекулярно-генетических данных, а на статистическом анализе возможного действия эпидемий оспы. «ГИПОТЕЗА КЛИМАТА». Кроме гипотез, связанных с инфекционными заболеваниями, предпринимались и другие попытки объяснить закономерности в пространственном распространении мутантного аллеля. Было изучено возможное действие климата и обнаружена сильная отрицательная корреляция между частотой мутации и рядом температурных факторов [Balanovsky et al., 2001, 2005; Limborska et al., 2002]. «ГИПОТЕЗА ВИКИНГОВ» постулирует, что мутация возникла в Скандинавии и распространилась по Европе в «век викингов», когда норманнские корабли достигали самых отдалённых побережий Европы [Lucotte, 2001]. ЕВРАЗИЙСКАЯ КАРТА РАСПРОСТРАНЕНИЯ МУТАЦИИ В целом надо признать, что факторы, повлиявшие на распространение мутации del32, поняты не до конца [Galvani, Novembre, 2005]. Для их лучшего понимания необходимо, в первую очередь, иметь точную картину распространения мутации в населении мира. Закономерности распространения мутации CCR5del32 в глобальном масштабе рассматривались как нами, так и многими иными авторами в целом ряде работ - по мере того, как накапливались данные о частотах мутации [Martinson et al, 1997; Libert et al, 1998; Stephens et al, 1998; Lucotte, 2001; Limborska at al, 2002; Galvani, Slatkin, 2003]. Недавно, объединив наши данные по русским и иным популяциям Восточной Европы с данными из опубликованных источников, мы составили общемировую сводку данных по частотам мутации del32. Эта база данных представлена здесь в виде карты распространения мутации в Евразии (рис. 6.1.1). Карта построена по данным о 185 популяциях, из них 35 - собственные неопубликованные данные авторов и их коллег [Balanovsky et al, 2005]. На карте {рис. 6.1.1.) видна четкая закономерность: частота максимальна на севере Европы (на побережьях Балтийского и Белого морей), и во всех направлениях от этой зоны частота плавно снижается. Модификация этой карты, вычленяющая основной паттерн изменчивости, демонстрирует концентрические круги - постепенное убывание частоты по мере удаления от зоны максимальных частот в Беломоро-Балтийском регионе [Balanovsky et al., 2005]. ИЗМЕНЧИВОСТЬ В РУССКОМ АРЕАЛЕ Для данного маркёра в масштабе Евразии свойственна едва ли не самая элегантная, четкая и простая пространственная изменчивость, которую только приходилось видеть авторам этой книги. Тем удивительнее, что распределение мутации в русском ареале носит довольно сложный характер (рис. 6.1.1). Наибольшие частоты отмечены на Русском Севере - во всех трёх изученных популяциях Архангельской области. Именно они формируют беломорскую часть беломоро-балтийского максимума, выявившегося при анализе в глобальном масштабе. К югу и западу русского ареала частота снижается. Таким образом, распределение мутации в русском ареале согласуется с глобальным трендом в двух аспектах: высокая частота на севере (в зоне беломоро-балтийского максимума) и снижение частоты к югу. РУССКАЯ АНОМАЛИЯ Но есть и важное расхождение. В глобальном масштабе частота мутации падаёт не только и не столько к югу, сколько с запада на восток - по мере удаления от Балтийского побережья. Но в русском ареале частота, напротив, снижается не к востоку, а к западу! Западные русские популяции (псковские, вплотную примыкающие к зоне Балтийского максимума, и смоленская популяция) обладают существенно меньшей частотой мутации (5-9%), чем центральные и даже восточные русские популяции (10-15%). Кроме того, Вологодская популяция, расположенная вблизи северного русского максимума частот (16-19%), имеет частоту мутации лишь 5%. Достоверность этой геногеографической аномалии подтверждается объёмами и качеством выборок. Трудно предположить, что могло сформировать такую чересполосицу в расположении зон высоких и низких частот в русском ареале. Эта аномалия достаточно ярко видна не только в «русском масштабе», но и является одним из важнейших нарушений плавного хода изолиний и на «евразийской» карте распространения мутантного аллеля {рис. 6.1.1). Остается надеяться, что по мере дальнейшего изучения «смоленско-псковско-вологодская» аномалия - зона низких частот, столь некстати расположившаяся рядом с зоной максимальных частот, будет более чётко оконтурена, что, в свою очередь, поможет лучше уяснить причины этого феномена. ВТОРОЙ МИРОВОЙ МАКСИМУМ Карта распространения мутации в Евразии (рис. 6.1.1), благодаря нашим новым (пока ещё неопубликованным) данным, выявила ещё один феномен - второй мировой максимум CCR5del32 в Западной Сибири. Пока он основан только на данных по шорцам (q=0.17). Данные по шорцам достаточно репрезентативны (154 человека) и представляют не одну, а несколько выборок с большой части этнического ареала шорцев. Аналогичные данные для шорцев были получены М. И. Воеводой с соавторами. В одной из популяций хантов [Yudin et al., 1998] выявлена высокая частота мутации (q=0.12; выборка 61 человек). Поэтому, хотя результаты требуют развертывания исследований в этом регионе, сам выявленный феномен второго мирового максимума может считаться установленным. ИТОГ Изменчивость CCR5del32 позволяет сделать два вывода в отношении русского генофонда. Во-первых, по этому маркёру выявилась особость населения Архангельской области. Причём, хотя эффективный популяционный размер именно на этих территориях особенно невелик, необычно высокие частоты нельзя объяснить лишь генетическим дрейфом. Ведь все три Архангельские популяции, причём географически и генетически изолированные друг от друга, отклонились от среднерусской частоты в сторону увеличения частоты мутации и примерно на одну и ту же величину, тогда как действие дрейфа предполагает случайное по направлению и силе отклонение. Во-вторых, изменчивость частоты этого маркёра в русских популяциях столь велика, что заметна даже в глобальном масштабе, на «евразийской» карте {рис. 6.1.1). 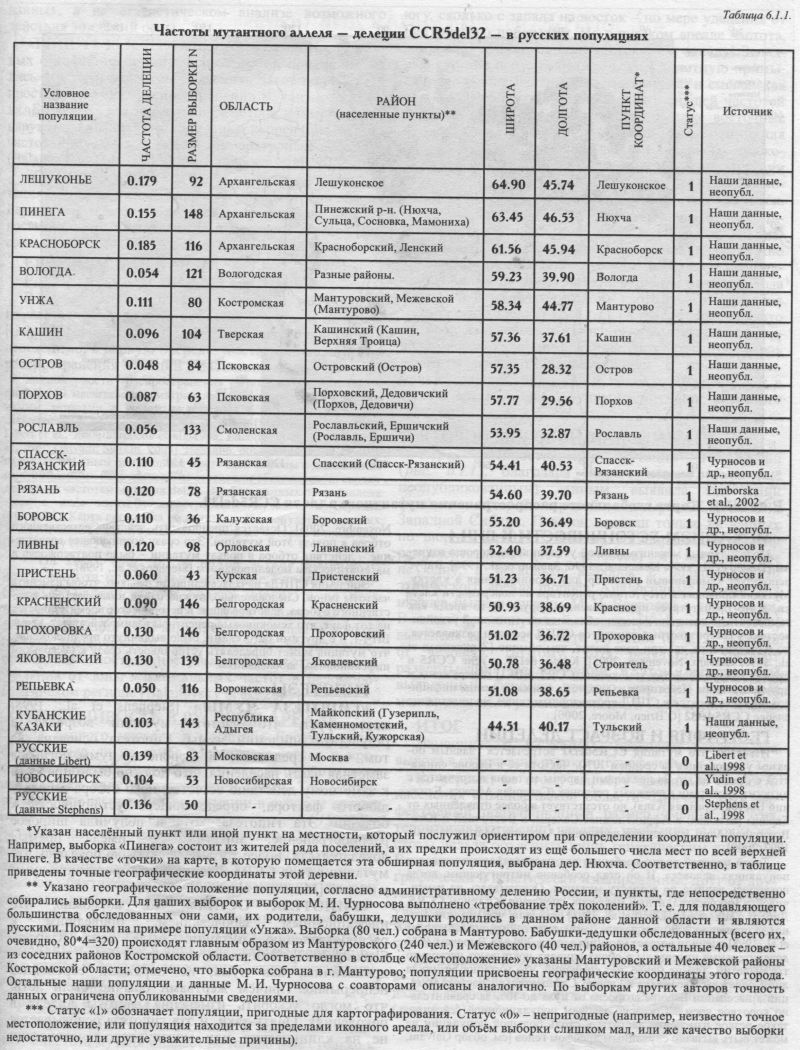
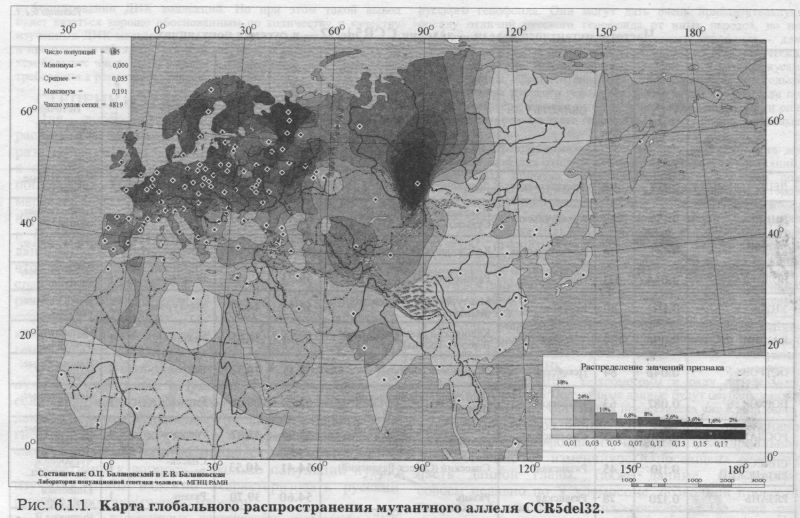
Рис. 6.1.1. Карта глобального распространения мутантного аллеля CCR5del32. |
загрузка...