1. Понятия
Многие результаты, представленные в этой книге, кажутся кроликом, вытащенным из шляпы фокусника. Чтобы не казалось, что они появились «ниоткуда» и «сами по себе», мы решили привести в некоторые основы геногеографии - ведь учебника по ней нет. В главе 1 мы рассмотрели лишь некоторые идеи и понятия, на которые опирается геногеография. Здесь, в Приложении, постараемся бегло обрисовать важнейшие черты этой науки. Основные понятия и инструменты пришли из популяционной генетики человека. Но все они непременно преломляются в свете географии - «субкультуры, озабоченной пространством» [Родоман, 1990].
§1. Ген и генофондГЕН И ГЕНОФОНД: ЧАСТНОЕ И ОБЩЕЕ В результате генных мутаций ген может принимать множество состояний. Каждое такое состояние называется «аллель». Представить себе генофонд можно как совокупность аллелей, циркулирующих в популяции. Но важнее не статическое определение генофонда, а способ его изучения. Популяционная генетика, изучая генофонд, прежде всего интересуется его разнообразием, его дифференциацией. Размах генетических различий между популяциями зависит от двух причин. Во-первых, от того, как долго и насколько разобщены популяции. Во-вторых, в какой степени и каким образом ген участвует в приспособлении популяции к особенностям среды (иными словами - от действия отбора). По приспособительному значению не только разные гены, но даже аллели одного гена могут резко отличаться друг от друга. А вот в том, что касается степени исторической разобщённости популяций, - здесь все гены находятся в равных условиях. Поэтому, если мы возьмём некий интегральный показатель по всем генам, и тем самым избавимся от их неравного приспособительного значения (от эффектов отбора), то с помощью этого показателя мы сможем увидеть, как генофонд зависит от истории популяции. Структура генофонда - это те реально существующие и наиболее общие закономерности, которые характерны для всего генофонда в целом. Поэтому в описание структуры генофонда не входит специфика каждого из генов. Изменчивость одного гена - это лишь одна из бескрайнего множества проекций многомерного строения генофонда. Ее описание дает столь частное видение структуры генофонда, что само по себе не способствует решению основной задачи геногеографии. Чтобы решить эту задачу, нужно изучить изменчивость одного за другим многих генов, а затем тем или иным способом обобщить эти данные, избавиться от случайностей и искажений «частного» видения, получить общие характеристики генофонда. Ясное понимание соотношения изменчивости отдельных генов и генофонда в целом - ключевой момент в геногеографии. Реально существующая, но недоступная непосредственному наблюдению структура генофонда проецируется во множество картин изменчивости, которые мы наблюдаем, перебирая один ген за другим. Отдельный ген выступает лишь в роли одного из маркёров генофонда, лишь как одно из бесконечного множества разнообразных и мелькающих отражений генофонда. Структура генофонда задаёт «общую модель» изменчивости каждого отдельного гена. Но оставляет ему значительную свободу «частного» проявления - картина изменчивости гена может значительно варьировать вокруг «общей модели» - по воле случая, под влиянием некоторых миграций или же по воле отбора. Поэтому так важно для изучения генофонда вычленить из изменчивости многих маркёров ту их общую составляющую, которая обусловлена общей структурой генофонда. СКОЛЬКО ГЕНОВ И КАКИХ?Здесь неизбежно возникает вопрос: какова количественная основа этой принципиальной разницы между двумя типами исследований? Если изучение общей изменчивости 120 маркёров считается изучением генофонда в целом, то будет ли достаточным изучение, например, 60 маркёров? Или 30? Или 10? Этот вопрос в геногеографии формулируется так: сколько и каких маркёров надо включить в анализ, чтобы, обобщив изменчивость этих отдельных маркёров, получить надёжные характеристики генофонда в целом? Необходимо убедиться, что разнообразие спектра маркёров отражает реальные характеристики генофонда. В таком случае по данным о нескольких маркёрах можно делать обоснованные выводы о генофонде в целом. Чем лучше в количественном и качественном отношении используемый набор маркёров, тем в большей степени его обобщённые характеристики приближаются к реальным характеристикам генофонда в целом. Действительно «полигенные» исследования с высокой точностью описывают реальные характеристики генофонда, поэтому их результаты могут обладать предсказательной силой. Примером могут служить главные компоненты генофонда Европы («полигенное» исследование) [Cavalli-Sforza et al„ 1994]: появляющиеся новые ДНК данные обязательно сравнивались с этим результатом, полученным по классическим маркёрам генетики - именно потому, что «старые» данные по классическим маркёрам рассматривались как прогноз для новых данных по ДНК маркёрам. Как известно, основной вывод, сделанный Кавалли-Сфорца с соавторами [Cavalli-Sforza et al., 1994] на основе изучения главных компонент народов Европы, это вывод о том, что ведущие черты генофонда Европы сформировались в неолите в ходе постепенного распространения земледельческого населения из Малой Азии через Балканы далее к северу и западу Европы. Результаты же работ последних лет, выполненных в большинстве случаев на маркёрах митохондриальной ДНК, свидетельствуют скорее о более раннем, чем неолитическая эпоха, времени формирования основных черт европейского генофонда. И может сложиться поверхностное впечатление, что новые работы по ДНК маркёрам опровергают старое исследование, выполненное по классическим маркёрам, что оно не отражало закономерностей генофонда в целом. И можно даже перейти от этого впечатления к мнению, что каждый новый тип маркёров будет по-новому описывать европейский генофонд, и что реальной структуры генофонда в действительности нет. Однако это впечатление обманчиво. В действительности, данные о «палеолитическом» времени возникновения присущих европейскому генофонду линий мтДНК могут вступать в противоречие только с «неолитической» интерпретацией главных компонент. Сам же факт основного направления изменчивости с юго-востока, установленный по классическим маркёрам, остаётся ни в коей мере не опровергнутым. Более того, он был подтверждён уже самыми первыми исследованиями по изменчивости ДНК маркёров в Европе. §2. Ареал популяцииСТРУКТУРА АРЕАЛА Для популяций человека важнейшими атрибутами являются брачная структура (доля браков, заключаемых вне популяции, или же степень нарушения свободы браков в пределах популяции) и ареал популяции. Ареал представляет собой жизненное пространство популяции и является важным фактором в её эволюции: ареал либо создаёт условия для формирования генетических различий, либо их фиксирует, если они возникли за счет иных причин. Оказывается, даже генофонд Москвы, где массы населения перемешиваются на клочке земли, не только в прошлом, но и сейчас обладает географической подразделённостью [Курбатова, 2004]. Тем более она была характерна для сельского русского населения, где брачные традиции («Хоть за курицу, но на свою улицу») приводили к генетическим различиям между популяциями. Даже для тех этносов, целостность которых поддерживается только культурной традицией (например, для евреев и цыган с их «виртуальными» ареалами), пространство остаётся важным источником генетических различий: географически разнесенные популяции по своим частотам генов зачастую сближаются с популяциями тех народов, среди которых они расселены [Mourant et al., 1978; Carmeli, Cavalli-Sforza, 1979; Karlin et al., 1979; Livshits et al., 1991]. Обычно, аргументируя несущественность ареала для этноса, приводят в пример устойчивые этносы с разорванными ареалами - например диаспоры и колонии. Однако с точки зрения генетики здесь проблемы, на наш взгляд, вообще нет. Если группа людей, проживающая вне основного ареала, придерживается прежней брачной структуры, то изменений в популяции и её генофонде просто не произошло. Неважно, где в тот или иной момент находится диаспора или колония - её члены в этом случае все равно генетически относятся к прежней популяции и участвуют в её воспроизведении. Можно считать, что её гены просто посланы «в командировку». Иной случай, если брачная структура резко меняется. Тогда независимо от того, заключают ли члены колонии только браки между собой в пределах своего нового ареала или же обмениваются генами с окружающими их популяциями, происходит формирование новой популяции с её собственным ареалом. Останется ли эта дочерняя популяция в рамках прежнего «материнского» этноса, или станет частью «удочерившего» её этноса, или же вообще станет со временем новым этносом - это дело не генетики, а истории и этнического самосознания. Генетика не определяет эти процессы, она лишь следует за ними. Для нее важно, что даже в «исключительных» этносах неоднородность популяции обычно сопряжена с географической подразделённостью. Тем более такая сопряжённость между подразделённостью самой популяции и структурированностью её ареала характерна для популяций, имеющих целостные, исторически сложившиеся и географически очерченные ареалы. Структурированность и целостность ареала вовсе не исключают, а обычно сопутствуют друг другу. Популяция населения крупного региона состоит из множества популяций разных иерархических уровней, вложенных друг в друга, как матрёшка в матрёшку. И ареалы этих субпопуляций также следуют «принципу матрешки». Структура и процесс формирования ареала часто является одной из важнейших страниц истории генофонда. Наиболее информативны эти страницы для тех народов, в жизни которых играли важную роль перемещения - массовые миграции, переселения, экспансия, колонизация новых территорий. История народа, история его генофонда порою может быть реконструирована через историю формирования ареала, который - при благоприятном стечении исторических обстоятельств - может сохранить память о развитии популяции не только в пространстве, но и во времени. МЕХАНИЗМЫ ФОРМИРОВАНИЯ АРЕАЛА Для популяций человека исторически сформировавшийся ареал популяции освящён памятью предков: любовь к отечеству - это зачастую «любовь к святым гробам»; это те корни, без которых человек становится «перекати-поле». Традиции, фольклор, предания и память народная любовно хранят черты и образы популяционного ареала, тех мест, в которых формировалась и жила популяция. Недаром столько крови проливается за «пядь родной земли», недаром поколения изгнанных народов живут мечтой о возвращении на свою историческую родину, на «свою» землю. И возвращаются, преодолевая немыслимые преграды, восстанавливая при этом не только ареал, но отчасти и генетическую структуру популяций. Наша многострадальная страна полна примеров этому. При этом ареал некорректно представлять лишь в двумерном пространстве географической карты. Популяционный ареал, как правило, содержит и иные измерения. Это может быть не только третье измерение физического пространства - высотное. В качестве иных измерений могут выступать любые факторы культуры (конфессиональные, лингвистические, этнографические и т. д.) или же факторы среды [Fisher, 1930; Edwards, Cavalli-Sforza, 1972; Алексеева, 1972, 1975, 1977, 1986, 1998; Дубинин и др., 1976; Sokal, 1979а; Бунак, 1980; Piazza et al., 1981а; Алтухов, Курбатова, 1984, 1990; Вавилов, 1987; Новорадовский и др., 1992; Спицын и др., 1985, 1994; Курбатова, 1996]. Например, в характеристике хозяйственно-культурных типов (степные номады, охотники-оленеводы, охотники на морского зверя и т. д.), звучит то дополнительное измерение ареала, которое свойственно данной популяции и обуславливает устойчивость ареала. Именно благодаря другим измерениям ареала возможны, например, случаи длительного мирного сосуществования в одном географическом пространстве двух популяций, ориентирующихся на разные природные ресурсы, или же принадлежащих к разным конфессиям, или относящихся к разным социальным слоям (сословия, касты). Важнейшим измерением при формировании ареала популяции являются поведенческие механизмы. Именно они чаще всего служат основой для возникновения симпатрического видообразования и существования симпатрических популяций [Dobzhansky, 1970; Майр, 1974; Яблоков, Юсуфов, 1976; Шмальгаузен, 1983; Воронцов, 1984; Северцов, 1987; Яблоков, 1987; Кайданов, 1996]. Если для птиц это может быть вариация песни, для грызунов - особенность их песенки-писка, для любых видов - своеобразие брачного поведения, то для человека поведенческие особенности, как правило, опосредованы через особенности культуры, воплощены в них. Весь исторически сложившийся облик обрядности, одежды, норм морали, пищи, домостроительства, множества этнографических, лингвистических, конфессиональных особенностей создают географически невидимые границы популяций. Это приводит к тому, что на одной территории определенное время могут существовать популяции, практически не заключающие между собой браки. Уникальным примером такой симпатрии, например, может служить изолят адыгейцев, древнего коренного населения Северного Кавказа. Их небольшая группа (причерноморские шапсуги) проживает среди массы пришлого населения на побережье Черного моря (от Туапсе до Сочи). Но даже в условиях огромного миграционного давления курортной зоны шапсуги сохраняют давние брачные традиции - подавляющая часть браков заключается в пределах популяций шапсугов общей численностью всего лишь 5 тыс. человек [Почешхова, 1998; Почешхова и др., 1998; Балановская и др., 1999]. АРЕАЛ В ГЕНОГЕОГРАФИИ Изучая популяции стандартными методами статистики, исследователь, как правило, стремится вычленить в наблюдаемой генетической изменчивости различия между популяциями (GST, FST, генетические расстояния). Но при этом неотъемлемое свойство популяции - её ареал - обычно так и остаётся за рамками исследования. Он не поддаётся обычному генетико-статистическому анализу - при кластеризации популяций и построении графиков главных компонент географическое положение популяций как бы исчезает из анализа. И потому полнота картины генофонда не достигается, а из межпопуляционной изменчивости исчезает её географическая суть. Пространство при этом служит своего рода «учётным бланком», в котором фиксируются точки — популяции. Ареалы популяций, их положение в пространстве не учитываются. И изучение межпопуляционной изменчивости сводится к сравнению чисел в точках, чисел в таблице. Географическая структура генофонда при этом остаётся неизвестной. Вместе с географией генофонда и часть его истории остаётся за рамками исследования. В географической структуре генофонда запечатлена динамика его формирования, поэтому географический подход позволяет через анализ пространственной изменчивости прийти к анализу истории генофонда. Но для этого пространство из фона должно стать организующим фактором. Это позволяет сделать лишь карта - лишь благодаря ей ареал становится важнейшим действующим лицом. Поэтому геногеография последовала за географией, сделавшей карту основой и инструментом тематических исследований. §3. Дрейф и миграции, мутации и отборГЕНЕТИЧЕСКИ ЭФФЕКТИВНЫЙ РАЗМЕР ПОПУЛЯЦИИ. Ne В популяции есть дети, взрослые, старики, и каждый из них несет два аллеля каждого аутосомного гена. Тогда, казалось бы, генетический размер популяций - то есть общее число генов в ней - можно просто считать в два раза большим, чем число людей в популяции. Но для изучения популяции важно знать, сколько генов будет передано следующему поколению. Следовательно, в подсчёт уже нельзя включать гены стариков и детей - генетического прошлого и генетического будущего популяции. Однако и оставшиеся гены также различны по своим судьбам. То, что взрослые, находящиеся в репродуктивном возрасте, оставляют разное число потомков, с точки зрения генетики означает, что они передадут меньшее или большее число копий своих генов следующему поколению. Чем больше в данной популяции семьи различаются по своему размеру, тем меньше генетически эффективный размер популяции. Но и это ещё не все: необходимо, чтобы дети - носители родительских генов - выжили, выросли, обзавелись своими семьями, оставили потомков, то есть, чтобы не прервалась передача генов по цепи поколений. Например, даже неравное соотношение полов в популяции может означать, что гены не всех мужчин и женщин будут переданы следующему поколению. И это немаловажно. Представим себе «гаремную» популяцию (в которой лишь малая часть всех мужчин передаёт свои гены следующему поколению), и пусть в каждом из 10 гаремов по 50 жен (итого, 510 человек, передающих свои гены следующему поколению). Генетически эффективный размер Ne такой популяции будет менее 40, а вовсе не 500, как было бы, если бы равное число мужчин и женщин (по 250) участвовали в передаче генов следующему поколению. Иными словами, генетически эффективный размер Ne нашей «гаремной» популяции с 510 репродуктивно активными членами окажется меньше генетически эффективного размера крошечной популяции с 20 мужчинами и 20 женщинами, но заключающими браки свободно и равноправно. Все эти и многие другие обстоятельства интегрированы в показателе генетически эффективного размера популяции Ne Обычно он составляет около 30% от общего числа генов в популяции. Это значит, что из всех генов популяции на каждый момент времени лишь треть связана с формированием генофонда следующего поколения. И генетические свойства нового поколения начинают зависеть от того, насколько полно (или же неравномерно) были представлены в этой трети гены родительского поколения. При оценке дифференциации генофонда через равенство Fe=1/(1+4NeMe) по умолчанию предполагается стационарность генетического процесса и постоянство эффективного размера популяций Ne, причём не только в пространстве, но и во времени. Этот вопрос рассматривался, в частности, в работах [Rogers, Jorde, 1995; Bowcock et al., 1991, Rychkov, Sheremetyeva, 1977], где показано, что для широкого круга популяционно-генетических задач эффективный размер популяций можно принять постоянным для всей ойкумены на протяжении последних 10 тыс. лет [Rogers, Jorde, 1995, Bowcock et al., 1991]. Это связано с тем, что в ряду популяций в пространстве, как и во временном ряду поколений одной популяции, генетически эффективный размер Ne определяется не как простая арифметическая средняя (∑Nk/K) по к популяциям, а как гармоническая средняя (1/к∑1/Nк)!. Так, например, для шести популяций с численностью 10, 100, Г000, 10Ό00, 100Ό00, 1Ό00Ό00, средняя гармоническая величина Ne будет равна лишь 50, а не 185 тысячам, как было бы в случае арифметической средней. Иными словами, генетически эффективный размер задаётся самыми «малыми» популяциями (как в примере с неравенством полов в «гаремной» популяции он задавался наименьшей - мужской - частью популяции). Очень важно и то, что рост численности населения главным образом связан с увеличением числа популяций, а не их среднего генетически эффективного размера. СЛУЧАЙНЫЙ ДРЕЙФ ГЕНОВ Ясно, что когда такая треть абсолютно мала (т. е. в нее входят «считанные» гены), случайные события легко могут исказить популяционную частоту гена при передаче из поколения в поколение. И чем меньше Ne, тем больше у потомков непредсказуемые отклонения от родительского генофонда. Когда такие отклонения накапливаются в долгой цепи поколений, то крайние звенья этой цепи становятся почти не похожими друг на друга. Процесс случайных отклонений в частотах генов, происходящих при передаче из поколения в поколение, официально назван «случайным дрейфом» генов. Это название удачно тем, что оно сразу говорит нам и о том, что изменения частоты гена в каждом поколении происходят чисто случайным образом (частота «дрейфует», как льдина); и о том, что в результате таких чисто случайных отклонений, популяции все же постепенно удаляются друг от друга. Они как осколки льдины, «отдрейфовывают» и друг от друга, и от материнской льдины, от которой они откололись - от прапопуляции. Если когда-то в результате событий истории две популяции разделились, а затем под действием дрейфа генов генетически удалились друг от друга, то генетическое расстояние между ними (расстояние, на которое они отдрейфовали) будет зависеть от двух величин - от генетического размера популяций (Ne) и от времени их раздельного существования (t). Значит, в генных различиях между популяциями в скрытом виде присутствует время, и время это измеряется поколениями. Как представить себе дрейф генов? Предположим, что исходная частота аллеля равнялась 0.5. Тогда это можно представить как огромный кувшин с разноцветными горошинами, где половина горошин - красная. Эта частота сохранится и в следующем поколении, если Ne велико - то есть если мы почти все горошины пересыпаем в следующий кувшин, в следующее поколение. Если же Ne мало, и мы зачерпываем лишь пригоршню горошин из генофонда, то случайно может оказаться, что в ней нет красных горошин (частота аллеля равна нулю), или напротив, все горошины красные (частота аллеля равна единице). Предположим, что мы изучаем не исключение из правил, не панмиксный мегаполис, а обычную небольшую популяцию, из которых и состояло человечество на протяжении тысячелетий, на протяжении почти всей своей истории. В ней из огромного множества гамет, произведённых всеми мужчинами и женщинами, в следующее поколение передаётся лишь горстка. Значит, в следующем поколении частота аллеля будет уже не 0.5, а чисто случайно изменится в ту или иную сторону. Итак, дрейф генов - это ошибка выборки из генофонда, совершаемая не исследователем, а самой историей. Ошибка тем больше, чем меньше генетически эффективный размер Ne - та часть популяции, которая должна передать генофонд следующему поколению. Чем меньше размер популяции, тем больше ошибка, тем сильнее случайный разброс в частотах гена от поколения к поколению, тем мощнее дрейф генов. ГЕНЕТИЧЕСКИ ЭФФЕКТИВНЫЕ МИГРАЦИИ Μe Обмен генами между популяциями называют миграцией генов независимо от того, что привело к проникновению «чужих» генов в популяцию - веками устоявшаяся структура брачных связей между соседними популяциями («поток генов»), случайные браки с пришельцами из других популяций или же перемещения целых групп населения. Количественной характеристикой этого процесса служит скорость притока генов за поколение, обозначаемая Мe. При этом учитывают не только общее число «прибывших» генов, но и степень их «новизны», несхожести с собственными генами популяции - чем «неожиданней» прибывший с миграцией вариант гена, тем больше генетическая эффективность миграции. При этом «новизна» прибывшего гена может определяться не только географической, но и культурно-исторической разобщённостью популяций. Представим, что в русской глубинке поселилась небольшая группа приезжих эфиопов и столь же немногочисленная группа переселенцев-белорусов. Поначалу и к тем, и к другим будут относиться как к чужакам, но потом станут заключать с ними браки, и со временем обе приезжие группы растворятся в местном населении. Будущим популяционным генетикам будет куда легче обнаружить миграцию эфиопов, чем след переселения белорусов: за счет резкого отличия мигрантов и принимающей популяции генетический эффект миграции эфиопов будет намного сильнее. ДРЕЙФ И МИГРАЦИИ Миграция генов - это социальный в своей первопричине процесс, который предотвращает утрату генетического сходства между популяциями в результате дрейфа генов. Любая популяция, таким образом, вовлечена в оба противодействующих процесса — дрейфа генов (Ne) и миграций генов (Мe). Их устойчивое противодействие задаёт новый предел для генетического расхождения популяций FGST=Fe. Благодаря потоку генов, новый предел различий между популяциями Fe оказывается гораздо меньше единицы (как ожидалось в случае действия дрейфа генов в условиях полной изоляции), а конкретная величина предельного значения Fe определяется теми историческими событиями, которые предотвратили потерю (или привели к появлению) общих генов. Это справедливо не только для взаимодействия разных популяций, но и между частями одной популяции, особенно если она расселена на большой территории. МУТАЦИИ И ОТБОР В популяциях человека действуют все факторы генетической динамики, и лишь их удельный вес различен в разных популяциях и для разных генов. Кроме двух только что рассмотренных главных факторов микроэволюции (дрейфа и миграций генов), есть ещё два фактора - мутации и отбор. При решении главной задачи книги - изучения структуры русского генофонда - они играют второстепенную роль, и потому скажем о них лишь несколько слов. Генные мутации изменяют сами гены, в то время как остальные факторы меняют частоты генов. В качестве фактора популяционной динамики действие мутаций оказывается настолько слабее дрейфа генов и миграций, что на популяционном уровне его уловить очень сложно (см. рассмотрение этого вопроса, например, в работе [Динамика популяционных генофондов... , 2004]). Исключение составляют однородительские маркёры - митохондриальной ДНК и Υ хромосомы. Восстановление исторической последовательности их мутаций и создаёт уникальную возможность установления исторической последовательности миграционных процессов. Отбор, как и мутации, действует на каждый ген в отдельности и по-своему, в отличие от дрейфа и миграций, которые влияют на все гены сразу и одинаково. Собственно исторический (то есть общественно-исторический) и микроэволюционный (то есть естественно-исторический) процессы различаются не только по силе воздействия на генофонд человека, но и по характеру воздействия на его гены. В исторический процесс все гены населения вовлечены в равной степени независимо от их функции в организме. В микроэволюционный процесс гены вовлекаются «индивидуально» и лишь в меру своей значимости для адаптации популяции к условиям среды. Воздействие исторического фактора на генофонд математически строго моделируется как селективно-нейтральный процесс. Некоторые результаты его воздействия, например различия между популяциями, можно определить достаточно точно. Однако для селективно-значимых генов на действие исторических сил накладывается влияние эволюционной силы отбора. Это увеличивает или уменьшает уровень изменчивости данного селективно-значимого гена FST(i)) по сравнению с общим для всех генов селективно-нейтральным уровнем Fe. §4. Дрейф и миграции в подразделённой популяцииИтак, два главных фактора - дрейф генов и их миграции определяют скорость и итоги селективно-нейтральной микроэволюции генофонда. Чтобы понять, какой смысл популяционные генетики вкладывают в эти слова, лучше всего вместо формул рассмотреть несколько примеров. ТОЛЬКО ДРЕЙФ ГЕНОВ Перед Вами (рис. 1.1., приводится по [Алтухов, 1983]) компьютерная модель дрейфа генов - компьютерный эксперимент, проведённый Юрием Петровичем Алтуховым и его сотрудниками для панмиксных популяций. Так называют популяции, которые не подразделены внутри себя - все члены такой популяции имеют равную вероятность заключить брак друг с другом. На старте десять одинаковых панмиксных популяции с одной и той же частотой гена (0.5). Или лучше представить их маршруты как десять вариантов эволюции одной популяции - варианты изменения частоты от одной и той же стартовой частоты гена (0.5) в некой «прапопуляции». Мы видим, как от поколения к поколению резко меняется - «дрейфует» - частота гена в пределах от 0 до 1. На финише лишь в одном случае из десяти популяций сохранился полиморфизм. Все остальные популяции содержат либо только «красные горошины», либо навсегда потеряли их. Это означает, что из-за дрейфа генов панмиксные - бесструктурные - популяции теряют память о прошлом генофонда. Это значит, что по современным популяциям (состояние в конце графика) мы не сможем восстановить, каким же был генетический облик прагенофонда, облик исходной популяции. Это судьба панмиксных изолированных популяций. Они утрачивают генетическую память: многие «предковые» аллели исчезли, частоты других - непредсказуемо изменились. ДРЕЙФ И МИГРАЦИИ Но теперь изменим лишь один параметр популяции - рассмотрим не панмиксную. а подразделенную внутри себя популяцию (рис. 1.2., приводится по [Алтухов, 1983]). Все то же, но каждая популяция разделена на подгруппы с небольшими миграциями между ними. В результате устойчивость генофонда резко возросла. На финише все десять подразделённых популяций сохранили полиморфизм и в какой-то степени воспроизводят исходную частоту гена, хотя и с большим разбросом. Это означает, что подразделённые популяции намного устойчивее к действию дрейфа генов и эволюционируют намного медленнее: частоты генов в субпопуляциях от поколения к поколению «скачут», но при этом средняя частота в тотальной «супер» популяции (в которую они входят) остаётся на месте. Это означает, что такая подразделённая популяция благодаря только одной лишь её подразделённости - имеет серьёзные шансы сохранить исходное разнообразие, сохранить генетическую память. 
ЭКСПЕРИМЕНТАЛЬНЫЕ ПОПУЛЯЦИИ Можно возразить: жизнь сложнее компьютерной модели. Согласны. Поэтому перед вами прекрасный эксперимент на живом объекте - дрозофиле. На рис. 1.3. (приводится по [Алтухов, 1983]) представлены результаты известных экспериментальных работ Юрия Петровича Алтухова и Елены Юрьевны Победоносцевой. Мухи были расселены по отсекам популяционного ящика и обменивались генами точно так, как мы предположили в предыдущей компьютерной модели - равная численность, равновероятный обмен между отсеками, но основная часть скрещиваний происходит в отсеке. Мы видим, что в подразделённой популяции средняя частота гена (белые кружки) остаётся практически неизменной: в течение 80 поколений варьирует около исходной частоты 0.5: от 0.6 до 0.4. Подразделённая популяция хранит генетическую память. Другой ящик тех же мух, с той же численностью и в тех же условиях, но с полной свободой скрещивания, - моделирует панмиксную популяцию (рис. 1.З.). В ней - за то же время - частота гена упала от 0.6 до 0.1 (черные кружки). Панмиксная популяция не помнит о прошлом, она теряет генетическую память. 
ОТ НЕОЛИТА ДО СОВРЕМЕННОСТИ Можно возразить: жизнь людей сложнее жизни мух. Возможно. Обратимся к человеку. На человеке эксперименты проводит сама история. Человек для популяционной генетики является также очень интересным объектом - у него детально изучена изменчивость в пространстве и времени. Нарис. 1.4. (приводится по [Rychkov, Sheremetyeva, 1977]) представлена характеристика популяций неолита и современности. Анализ был проведён Юрием Григорьевичем Рычковым и Аллой Арменовной Мовсесян по дискретно варьирующим признакам черепа. Предполагается, что эти признаки (дополнительные швы, вставные косточки, отверстия) по высокой наследуемости приближаются к генетическим маркёрам. Частота каждого из 12 признаков отложена на соответствующем радиусе, и получен полигон - своеобразный схематический «портрет» популяции. Мы получаем уникальную возможность заглянуть в прошлое - ведь прямо по ДНК маркёрам такой анализ пока провести нельзя. Современный генофонд представлен 24 коренными народами Сибири. Неолит представлен шестью палеоантропологическими сериями, охватывающими довольно большой промежуток времени. В среднем современность и неолит разделяет более пяти тысяч лет, 200 поколений сменились за это время. Мы видим, как резко различаются портреты всех современных популяций и насколько не похожи друг на друга древние популяции. Сходными во всей серии,состоящей из 31 «портрета»,оказались лишь два«портрета»: обобщённый «портрет» современности и обобщённый «портрет» неолита. В данном виде анализа вся Сибирь рассматривается как одна подразделённая популяция. Прошли тысячелетия, а средние параметры этой подразделённой популяции все те же: неолит и современность отличаются друг от друга меньше, чем нынешние народы, живущие по соседству. Подразделённая популяция хранит генетическую память и проявляет фантастическую устойчивость своего генофонда. УСТОЙЧИВОСТЬ ПОДРАЗДЕЛЕННЫХ ПОПУЛЯЦИИ К сожалению, ДНК маркёры пока не позволяют разглядеть генофонды прошлого. Но можно привести и другие примеры - например, с использованием не дискретно варьирующих Сравнение распределений 12 независимых признаков в неолитических (А) и современных (В) популяциях Сибири показывает, что невзвешенные частоты qt в современной популяционной системе являются хорошими оценками q0 в предковой неолитической популяции, t = 200 поколений. В центре рисунка помещена схематическая диаграмма теоретического оправдания такого сравнения. Современные популяции: I - азиатские эскимосы; II - чукчи Чукотсткого полуострова; III - алеуты; IV - негидальцы бассейна Амура; V - ульчи нижнего Амура; VI - тунгусы Восточной Сибири; VII - буряты Прибайкалья; VIII - монголы; IX - тувинцы Саянского нагорье; X - теленгиты Алтая; XI - хакасы Минусинской котловины, Енисея; XII - селькупы Западной Сибири; XIII - манси восточных склонов Урала; XIV - ханты бассейна Оби. Неолитические популяции: а - серовская культура, Ангара; b - китойская культура, Ангара; с - глазковская культура, Ангара; d - серовская культура, верхняя Лена; е - глазковская культура, верхняя Лена; f- глазковская культура, Забайкалье. Частота q = 0 в центре круга; q = 0.3 на периметре. 1 -12: признаки, перечень которых приведен в [Рычков, Мовсесян, 1972; Мовсесян, 1973; Рычков, 1973]. признаков черепа, а фамилий. Полные родословные, собранные Эльвирой Аслановной Почешховой уникальны - они охватывают всех ныне живущих адыгов-шапсугов и уходят на глубину шести поколений. Мы вместе Э. А Почешховой и Ю. А. Серегиным провели анализ состава фамилий в каждом поколении, а затем построили обобщённые карты распространения фамилий для каждого поколения шапсугов. Они позволили своими глазами увидеть, как меняется генофонд в пространстве и времени (глава 10, рис. 10.4.3.). Оказалось, что все шесть карт чрезвычайно похожи. Коэффициент корреляции между самыми удалёнными во времени картами - первого и шестого поколений - невероятно высок: он составил r=0.85. Мы видим, что генофонд сохранил себя на протяжении всех шести поколений. Но самое удивительное в том, что это за народ. Мы видим отражение потерянного навсегда. Адыги-шапсуги - древнее население Северного Кавказа. Их сплошной ареал простирался от Кубани до Черного моря. Но их смела Кавказская война. К 1865 году все причерноморские аулы шапсугов были уничтожены, а прикубанские - переселены. Сейчас от них осталось два осколка: 5 тысяч человек в Прикубанье, и столько же - в предгорьях у Черного моря. Но они сохранили отражение разрушенного генофонда. У нас есть основания считать, что Прикубанье отражает прежнюю «Малую Шапсугию», жившую к северу от Кавказского хребта. А в Причерноморье постепенно просачивались те, кто остался в живых из «Большой Шапсугии». Они старались селиться именно в той долине, где испокон веков жили их предки. Они сохранили традиции брачной структуры. Подразделённая популяция вновь доказала нам свою необычайную устойчивость. 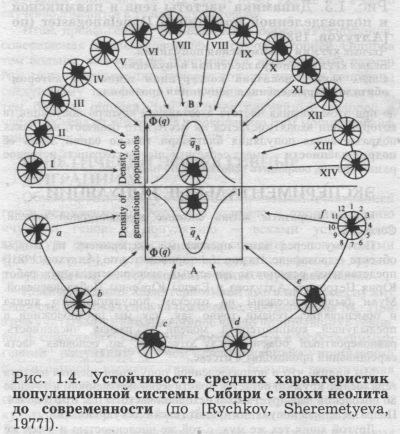
ИТОГИ МИКРОЭВОЛЮЦИИ Все приведённые примеры показывают, что подразделённая популяция устойчива и хранит генетическую память, а панмиксная - нет. Мы убедились, что микроэволюция в этих популяциях идет с разной скоростью и приводит к разным результатам. Разное соотношение дрейфа и миграций в ходе микроэволюции мы видим в её итогах - в пространственной изменчивости популяций - независимо от того, рассматриваем ли мы эти итоги с помощью карт или же оценок различий между популяциями FST≈1/(1+4NeMe). Например, и карты, и FST-статистики свидетельствуют о генетическом «однообразии» населения Европы и о генетическом разнообразии коренного населения Сибири. Отличия этих двух генофондов поражают: генетическое разнообразие локальных популяций в пределах одного «среднего» сибирского народа больше, чем все генетические различия между всеми народами Западной Европы! Столь впечатляюще разные итоги микроэволюции в Сибири и в Европе (глава 9, рис. 9.3.3.) заданы лишь разным соотношением дрейфа и миграций генов. Вот насколько мощны эти два фактора микроэволюции. §5. Подразделённость генофондаВ предшествующем разделе мы видели, как наличие в популяции подразделённости резко меняет её свойства: и дрейф генов, и миграции начинают действовать по-другому, а сама популяция приобретает генетическую устойчивость. Это, возможно, самый яркий, но далеко не единственный пример того огромного значения, которое имеет подразделённость популяции для её генофонда. Роль подразделённости столь велика, что её иногда (наряду с дрейфом генов, миграциями, мутациями и естественным отбором) называют ещё одним, пятым фактором микроэволюции. Это, конечно же, не вполне верно. Ведь фактор подразделённости влияет на генофонд не непосредственно, а через «обычные» факторы микроэволюции. Но это влияние столь ощутимо, что у самых разных авторов возникает желание упомянуть подразделённость (в виде популяционной структуры или же в обличии инбридинга) при перечислении важнейших факторов, определяющих генофонд. В классической популяционной генетике подразделённость популяции является одним из центральных моментов: на ней строится и теория инбридинга, и расчёт межпопуляционной изменчивости, и анализ иерархически организованных популяций. Именно подразделённости посвящены множество математических моделей и основополагающие работы Самуила Райта, Масатоши Нея и многих других «классических» популяционных генетиков. Мы не будем пытаться дать здесь сколько-нибудь полный обзор этого «главного» раздела популяционной генетики, а рассмотрим лишь несколько моментов, важных для анализа пространственной структуры русского генофонда. ЧТО ТАКОЕ ПОДРАЗДЕЛЁННАЯ ПОПУЛЯЦИЯ Чтобы представить, что генетики понимают под подразделённой популяцией, достаточно лишь знать, что её противоположностью является популяция панмиксная. В панмиксной популяции все члены имеют равную вероятность вступить в брак друг с другом - внутри панмиксной популяции нет барьеров для потока генов. А внутри подразделённой популяции такие барьеры есть. Подразделённая популяция - та, которая в процессе передачи информации в поколениях оказывается подразделённой на части: внутри каждой части поток генов свободен, а между частями -ограничен. Панмиксная популяция похожа на цельный листок березы или липы, а подразделённая - на рассечённый листок клевера. Как лист рябины состоит из нескольких мелких листочков, так и подразделённая популяция состоит из нескольких субпопуляций. Подразделённость, то есть наличие барьеров для потока генов, может быть реализована самыми разными способами. Самый очевидный и самый частый - это пространственная, географическая подразделённость: популяция, расселившись на большой территории, естественным образом распадается на части, соответствующие географическому членению этой территории. Например, народы, живущие в горах, часто подразделяются так, что каждая группа занимает свою долину или свое ущелье. А живущие на равнине русские тоже подразделяются, но уже не по долинам, а по географическим областям. Ведь очевидно, что рязанец скорее женится на живущей по соседству рязанке, чем на далекой неизвестной ему вологодской красавице. Именно таким образом пространство, ареал популяции становится тесным образом связанным со структурой популяции, с её подразделённостью на составные части. Подразделённость может быть реализована и множеством других способов. Например, обычай заключать браки среди кровных родственников тоже нарушает панмиксию. А значит, в популяции появляется некая, пусть слабая, подразделённость, которую в данном случае мы назовём неслучайным инбридингом. Неслучайным - потому что он происходит вследствие неслучайного, ассортативного скрещивания - родственники заключают браки преимущественно с родственниками, и это приводит к увеличению доли гомозигот в популяции. А сама популяция как бы подразделяется внутри себя на группы тех, кто является в большей степени родственниками. Обычай кровнородственных браков распространён, например, в Дагестане и среди многих других горных народов, где он вызван, как считается, экономическими причинами (стремлением сохранить самую большую ценность - с трудом созданный в горах надел земли - в руках одной большой семьи). Явление, совершенно отличное внешне, но равнозначное с точки зрения популяционной генетики, можно наблюдать на противоположном конце Кавказа. Небольшие, изолированные друг от друга популяции адыгейцев не допускают кровнородственные браки вплоть до седьмой степени родства. Но в силу самой изолированности аулов дрейф смещает частоты генов в каждом ауле независимо, и, рассматривая всю систему из нескольких аулов, мы опять-таки видим увеличение доли гомозигот (явление, известное в популяционной генетике как эффект Валунда или действие подразделённости). И потому это явление возрастания гомозиготности популяционный генетик тоже назовёт инбридингом - но, в отличие от дагестанского варианта, случайным инбридингом. ВЕЗДЕСУЩНОСТЬ ПОДРАЗДЕЛЁННОСТИ Тут мы сталкиваемся с очень важным моментом геногеографии. Возникает вопрос - а что нам считать популяцией? Большую подразделённую популяцию (всех русских) или её отдельные «разделы» (население Рязанской, Вологодской и других областей)? Что правильнее: считать популяцией отдельный аул адыгейцев, а всех адыгейцев считать «суперпопуляцией», или же адыгейцев считать «просто популяцией», а отдельные аулы - её частями? Для популяционной генетики этот вопрос не имеет смысла. Исследователь может выбирать для рассмотрения популяцию любого иерархического уровня - от небольшой индейской деревушки до населения всей Америки. Но что имеет огромный смысл и обязательно для каждого - это всегда, в любом случае отдавать себе отчет в том, популяцию какого уровня мы в данный момент рассматриваем. Все человечество представляет собой одну гигантскую популяцию, подразделённую на множество меньших, но все ещё огромных популяций, которые дальше подразделяются на всё более дробные. Это принцип матрёшек, с тем отличием, что в большую матрёшку входит только одна следующего размера, а популяция обычно включает множество вложенных в нее одинаково меньших «матрёшек». РАЗЛИЧИЯ МЕЖДУ ЧАСТЯМИ ПОДРАЗДЕЛЕННОЙ ПОПУЛЯЦИИ Легко видеть, что лист дуба подразделён на лопасти, но лист каштана или смоковницы подразделён намного больше. Популяции тоже различаются по степени подразделённости. Эту степень можно измерить несколькими способами. Во-первых, можно сравнить, насколько отдельные части (субпопуляции) генетически отличаются друг от друга. Для этого нужно лишь знать частоты генов в каждой из субпопуляций и вычислить величину генетических различий между ними. То же самое можно описать и чуть другими словами: чтобы определить степень подразделённости популяции, нужно рассчитать дисперсию частоты гена в субпопуляциях. Понятно, что если популяция в большой степени подразделена (субпопуляции в большой мере изолированы друг от друга), то частота гена в субпопуляциях значительно варьирует, и дисперсия частоты будет велика. Второй способ измерить подразделённость состоит в расчёте гетерозиготности. Как известно из правила Харди-Вайнберга, в панмиксной популяции с частотой аллеля q гетерозиготность составляет 2q(l-q). А в подразделённой популяции с той же средней частотой аллеля q гетерозиготность оказывается меньше (так называемый эффект Валунда). Причём это снижение гетерозиготности тем больше, чем выше степень подразделённости популяции. Поэтому, сравнивая среднюю гетерозиготность субпопуляций (обозначаемую1 HS) с теоретически ожидаемой гетерозиготно-стью тотальной популяции HT, можно вычислить степень подразделённости популяции, обозначаемой как GST. Формула расчёта очень проста: GST=(HT-HS)/HT Оба способа расчёта - и через дисперсию, и через снижение гетерозиготности - измеряют одну и ту же подразделённость. Поэтому, при условии использования адекватного математического аппарата, эти две величины оказываются равны друг другу. РАЗЛИЧИЯ МЕЖДУ ПОПУЛЯЦИЯМИ Существует много способов сравнить две популяции и определить степень их генетического различия. Эти меры получили название генетических расстояний (генетических дистанций). Все они сравнивают, насколько различаются частоты аллелей в двух популяциях. Возникает вопрос, как быть, если мы хотим сравнить не две, а три или большее число популяций? Для этого можно рассчитать генетические расстояния между каждой парой популяций. И увидеть, какие из популяций больше похожи друг на друга, а какие от них отличаются. Если же усреднить все попарные различия между популяциями, то мы получим как бы общую характеристику всей рассматриваемой системы популяций - насколько все входящие в систему популяции генетически отличаются друг от друга. Легко видеть, что по смыслу это та же самая степень подразделённости, которую мы описывали совсем недавно, только с помощью других мер различий. Для этого нужно лишь сменить точку зрения на ситуацию: вместо сравнения отдельных популяций, составляющих вместе «систему» популяций, начать говорить о различиях между субпопуляциями, входящими в подразделённую популяцию. Мы уже упоминали, что отличие «популяции» от «субпопуляции» заключается лишь в точке зрения: практически каждая популяция является звеном в цепи матрёшек - она состоит из нескольких популяций низшего уровня (субпопуляций), но и сама является субпопуляцией по отношению к группе населения более высокого иерархического уровня. Например, популяция «население Костромской области» состоит из субпопуляций - районов области, но в свою очередь является частью популяции «русский народ». Вернемся к различиям между группой популяций. Мы сказали, что, рассчитав генетические расстояния между каждой парой популяций и усреднив все эти попарные отличия, мы получим величину подразделённости для данной группы популяций. Приятная неожиданность заключается в том, что этот (третий уже!) способ рассчитать подразделённость даст нам опять то же самое число, ту же величину подразделённости, что и два другие способа расчёта (через дисперсию частоты аллеля в субпопуляциях и через снижение гетерозиготности). Поэтому разные меры генетических расстояний часто являются одновременно и мерами подразделённости. Например, часто используемый способ расчёта генетических дистанций - FST дистанции, и эта математическая величина является очень близким родственником классическому показателю подразделённости: FST-статистике Райта. Аналогично, неевские генетические расстояния (рассчитываемые по формуле, выведенной М. Неем) легко превращаются в неевскую же меру межпопуляционной изменчивости GST. Есть и четвертый способ расчёта подразделённости, и он крайне важен. Дело в том, что он напрямую связывает подразделённость популяции с генетическим дрейфом и миграциями. По-пуляционный смысл этой связи в том, что генетические различия между субпопуляциями возникают за счет дрейфа, но уменьшаются за счет миграций между субпопуляциями (§4). Подразделённость (Fc) рассчитывается по формуле Fe=1/(1+4NeMe), где Ne - генетически эффективный размер популяции, задающий интенсивность дрейфа, а Мe - генетически эффективные миграции (§3). ПОДРАЗДЕЛЕННОСТЬ. ДИФФЕРЕНЦИАЦИЯ И МЕЖПОПУЛЯЦИОННАЯ ИЗМЕНЧИВОСТЬ Остается добавить, что у понятий «подразделённость популяции» и «генетические расстояния между популяциями» есть ещё два синонима: это «дифференциация» популяционного генофонда и «межпопуляционная изменчивость». Все эти термины несколько с разных сторон описывают одно и то же явление - подразделённость человечества на множество иерархически соподчинённых (вложенных одна в другую) популяций. Мы видим, что все показатели, описывающие генофонд популяции, теснейшим образом связаны с понятием подразделённой популяции. Генетические расстояния превращаются в подразделённость, дисперсия частоты аллеля оказывается связанной с гетерозиготностью, и опять-таки измеряет степень подразделённости, и к той же величине подразделённости приводит соотношение дрейфа и миграций. Целью этого параграфа 5 было показать читателю, мало знакомому с теорией популяционной генетики, важность понятия подразделённой популяции и ввести термины, которыми мы широко пользуемся во всей книге. На взгляд авторов, важность концепции подразделённости вызвана именно тем, что все человечество представляет собой многоуровневую, иерархическую популяцию, и почти каждая конкретная популяция и сама состоит из нескольких субпопуляций, и входит в состав популяции более высокого иерархического уровня. В следующем разделе 2 приводятся некоторые конкретные сведения об используемых мерах подразделённости, а также описывается наиболее сложный случай - анализ многоуровневых популяционных систем, когда каждая субпопуляция в свою очередь состоит из нескольких «субсуб-популяций». 1 Подстрочные индексы имеют свой смысл: S обозначает субпопуляцию (Subpopulation), Т - тотальную популяцию (Total population), а индекс ST - соотношение субпопуляции и тотальной популяции. Например, гетерозиготность обозначается буквой Н. тогда HS обозначает гетерозиготность субпопуляции, НT - гетерозиготность тотальной популяции, a FST- изменчивость (F) субпопуляций в пределах тотальной популяции. |
загрузка...