8.2. «Главные сценарии»
|
§1. Классические маркёры: Зримый образ генофонда - Главный сценарий - Долготная изменчивость - Широкое поле встречи запада и востока - Европеоиды и уралоиды - Второй сценарий - Широтная изменчивость - Дыхание степей Евразии - Связь с климатом
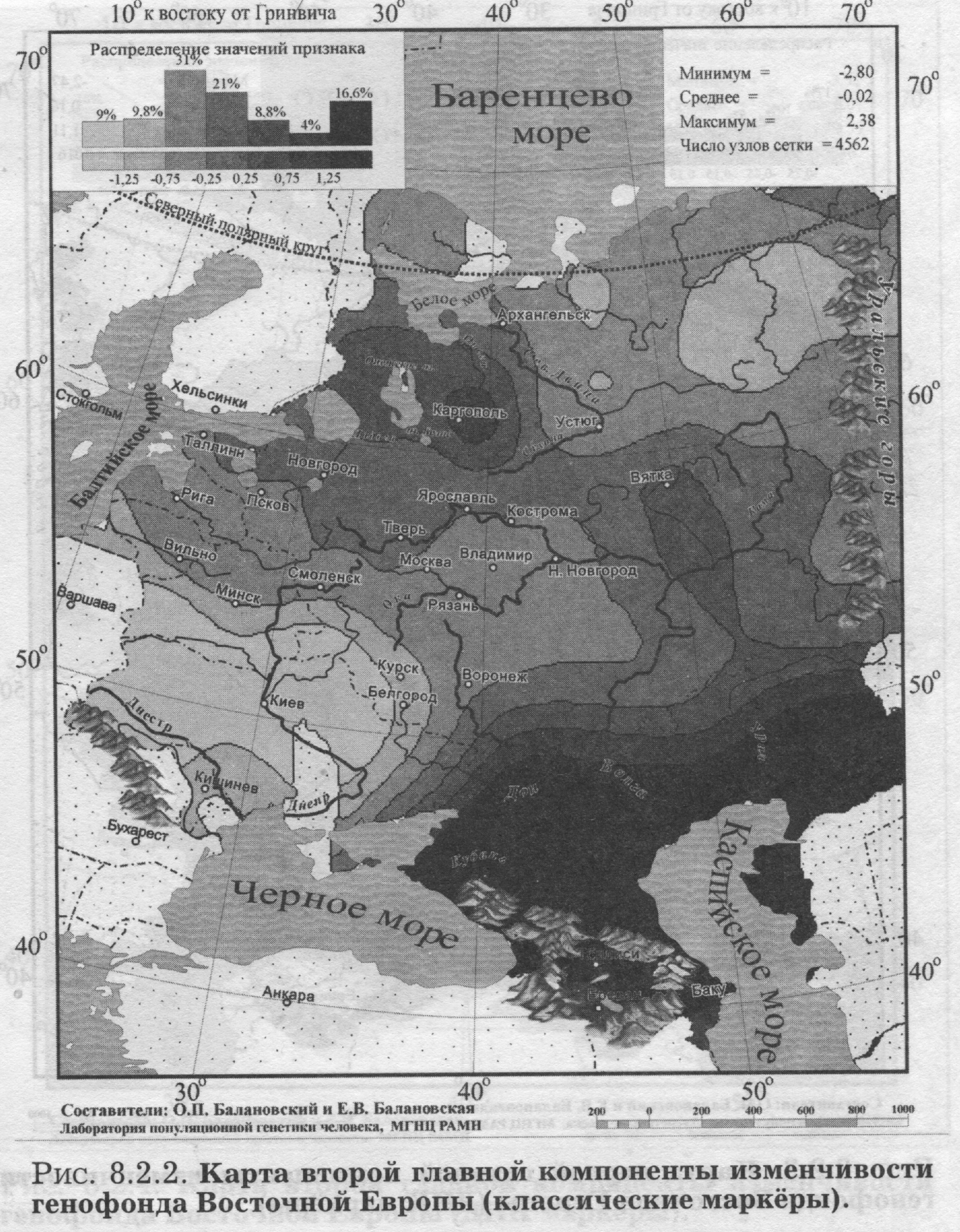
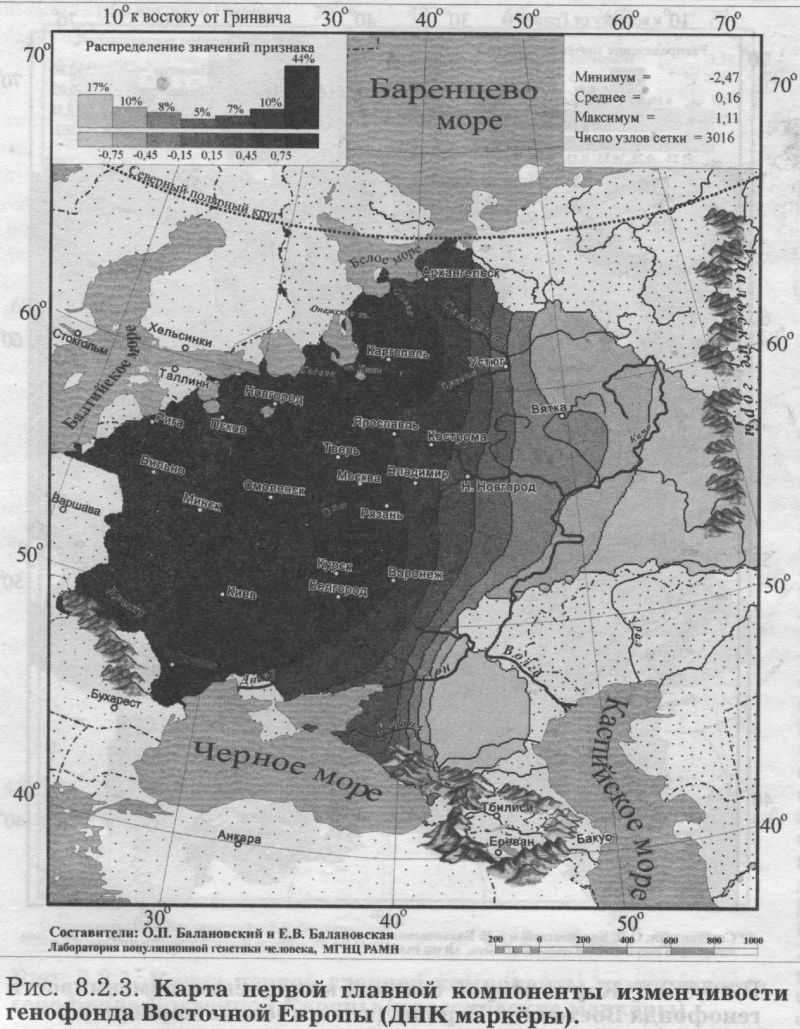
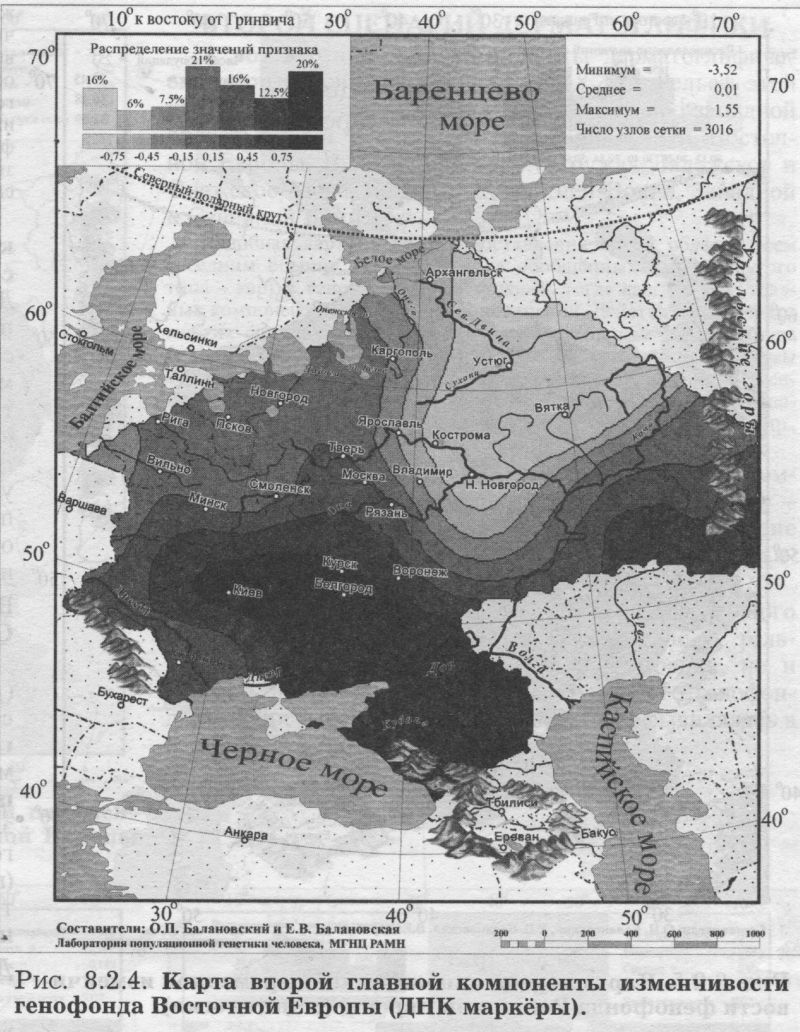
§2. Аутосомные ДНК маркёры: Главный сценарий - Вновь долготная изменчивость - Узкая полоса встречи запада и востока - Второй сценарий - Вновь широтная изменчивость - Финно-угорский след на северном экстремуме - Третий сценарий - Вновь сложная седловидная структура - Впервые показано сходство карт трёх главных сценариев обоих типов аутосомных маркёров - Важный теоретический вывод - Глубокое сходство картин мира по ДНК и по классическим маркёрам §3. Антропология - соматология и дерматоглифика: Большой и единый набор популяций - Признаки видимые и осязаемые - Соматология - Вновь долготная изменчивость на первом сценарии - Вновь седловидный рельеф на второстепенном сценарии - Дерматоглифика - Снова долготная изменчивость на первом сценарии - Второй сценарий необычен - Балтийский след §4. Митохондриальная ДНК: Тренды отдельных гаплогрупп - «Азиатские» гаплогруппы часты в Восточной Азии, убывают на запад к Уралу и почти не достигают восточных славян - Разнообразие гаплотипов снижается к северу - Первый сценарий - Опять долготная изменчивость - Славяне в кольце финно-угров - Кто в Европе самый разный? В предыдущем разделе мы провели статистический анализ генофонда народов Восточной Европы (с помощью GST и HS). Теперь можно перейти к изучению пространственной изменчивости того же генофонда с помощью обобщённых-компьютерных карт. Картографический анализ генофонда Восточной Европы проведём, опираясь на пять разных типов признаков. Вновь, как и в статистическом анализе, это будут классические и аутосомные ДНК маркёры, и ещё к ним добавятся данные соматологии, дерматоглифики и гаплогруппы митохондриальной ДНК. Такой «мультиокулярный» подход, использующий свидетельства пяти столь разных очевидцев, позволяет впервые создать наиболее полную, а потому максимально объективную, картину изменчивости восточноевропейского генофонда. §1. Классические маркёрыДЛЯ картографического анализа классических маркёров взяты те же локусы и аллели, что и для статистического анализа (табл. 8.1.1). Но на этот раз мы не отбирали этносы, сопоставимые с изученностью ДНК маркёров, а проанализировали всю имеющуюся информацию по классическим маркёрам (Приложение, разделы 5 и 6). Карты главных компонент изменчивости построены по наиболее изученным классическим маркёрам: 100 аллелей 33 локусов. Первая компонента описывает 19.7% общей дисперсии всех генов, вторая вбирает в себя 10.1%, третья - 7.6% общей дисперсии. Мы видим резкое падение дисперсии, указывающее на высокую эффективность метода главных компонент. Первая компонента выявляет пятую часть всей информации, содержащейся в исходном массиве данных - при неэффективности метода она составила бы лишь полтора процента3. Поэтому карта первой главной компоненты отражает основную закономерность в изменчивости классических маркёров в ареале Восточной Европы её главный сценарий. ПЕРВЫЙ СЦЕНАРИЙ Первая компонента генофонда изменчивости классических маркёров (рис. 8.2.1) характеризуется той плавностью и простотой, которая позволяет картографическому образу становиться зримым образом генофонда. Значения компоненты постепенно изменяются от юго-запада к востоку. Основное направление изменчивости - долготное: изолинии вторят меридианам, и корреляция с долготой составляет r=0.81.4 Карта выявляет две зоны, западную и восточную, плавно перетекающие друг в друга. Следуя интерпретации нашего предыдущего варианта этой карты, построенной по меньшему объему данных [Рычков, Балановская, 2005], можно думать, что здесь отражено взаимодействие двух пластов расового состава. Основанием служит совпадение западной зоны с ареалом европеоидных народов Восточной Европы, а восточной зоны - с ареалами народов с той или иной степенью монголоидной примеси [Рогинский, Левин, 1978]. Кроме того, долготное направление изменчивости также соответствует основному направлению смешений европеоидных и монголоидных пластов населения [Алексеева. 1999]. Но монголоидная составляющая в населении Восточной Европы представлена не классическими монголоидами [Хрисанфова. Перевозчиков, 1999], а народами уральской расы. Уральская же раса представляет собой своеобразное сочетание европеоидных и монголоидных особенностей, которое нельзя свести к механическому смешению двух больших - европеоидной и монголоидной - рас [Бунак. 1980; Рогинский Левин, 1978; Хрисанфова. Перевозчиков, 1999; Моисеев, 1999; Перевозчиков. 2005]. Учитывая это, следует трактовать карту первой главной компоненты как выявляющую уральскую. а не непосредственно монголоидную расовую составляющую. Корректнее здесь использовать классификацию В. В. Бунака на западный и восточный расовые стволы [Бунак, 1980]. Впрочем, эти интерпретации достаточно близки, поскольку все они утверждают взаимодействие европеоидного и «не европеоидного» пластов в составе восточноевропейского генофонда. Большая часть Восточной Европы занята промежуточными значениями главной компоненты. Такой плавный переход значений естествен для восточноевропейского региона, на территории которого смешение европеоидных и монголоидных (возможно, точнее говорить «уралоидных») народов происходило со времени формирования этих рас [Бунак, 1980; Алексеев, 1974]. ВТОРОЙ СЦЕНАРИЙ Вторая и третья главные компоненты всегда описывают меньшую часть информации, содержащейся в исходном массиве данных, и потому менее важны для изучения генофонда, чем первая компонента. Однако они необходимы для полноты картины: если первая компонента отражает основную закономерность генофонда, то вторая и третья компоненты - следующие по значимости, но тоже объективно существующие закономерности. На рис. 8.2.2. представлена карта второй главной компоненты по классическим маркёрам. Она выявляет изменчивость, близкую к клинальной: значения компоненты плавно меняются от юго-востока к северным и западным популяциям. Таким образом, вторая компонента классических маркёров (рис. 8.2.2.) определяется юго-восточным, прикаспийским экстремумом, занимающим предгорья и степи между Кавказом и Южным Уралом. Вся остальная (много большая) территория Восточной Европы противопоставляется этому юго-восточному полюсу. Такая изменчивость может отражать влияние генофонда степных народов (преимущественно алтайской языковой семьи). Многие миграции из внутренних районов Азии на территорию Восточной Европы шли по степной полосе и связаны именно с юго-востоком Европы (от Южного Урала до Северного Причерноморья). Представляется вполне вероятным, что эти миграции различных степных народов, направлявшихся примерно по одному маршруту - по степной зоне Евразии - в течение многих столетий (от гуннов в IV веке до калмыков в XVII) оказали значительное влияние на восточноевропейский генофонд. Можно предположить, что именно влияние степных народов (главным образом алтайской семьи) и сформировало тренд изменчивости генофонда с юга и юго-востока, который выявляет вторая главная компонента. Вместе с тем, возможна и иная трактовка карты второй главной компоненты: поскольку выявляемая тенденция достаточно близка к широтной изменчивости (коэффициент корреляции с широтой r=0.72), нельзя исключать возможность её связи с климатическими параметрами. Широтная изменчивость второй главной компоненты в значительной мере сходна с картиной широтной зональности в Восточной Европе, и в таком случае вторая главная компонента, возможно, отражает адаптацию генофонда к различным условиям среды. В настоящее время нет достаточных оснований, чтобы сделать выбор между двумя интерпретациями второй главной компоненты: экологической (связь с климатом) и исторической (влияние генофонда степных народов). Да и вряд ли возможно разделить историю и широтную зональность природы - ведь с экологическими зонами были связаны потоки исторических миграций, почему мы и говорим о взаимодействии населения «леса» и «степи». ТРЕТИЙ СЦЕНАРИЙ Карта третьей главной компоненты демонстрирует довольно сложную картину с четырьмя ядрами экстремумов. Её сходство с третьим сценарием аутосомных ДНК маркёров столь велико, что мы перенесём её описание в следующий параграф. §2. Аутосомные ДНК маркёрыАнализ главных компонент изменчивости ДНК маркёров в ареале Восточной Европы проведён для тех же шести локусов, что и статистический анализ (табл. 8.1.1). Были построены 114 карт распространения каждого из 114 аллелей этих ДНК локусов. Однако, большая часть аллелей микро- и минисателлитов в Восточной Европе очень редка - лишь для 51 аллеля средняя частота превысила планку 1%. Поэтому обобщённые карты построены максимально строго - на основе только тех 51 карт аллелей, которые удовлетворяют 1% критерию полиморфизма. ПЕРВЫЙ СЦЕНАРИЙ На карте первой главной компоненты (21% обшей дисперсии, рис. 8.2.3.) изменение значений географически чётко очерчено как долготное: изолинии в целом следуют меридианам, и основное направление изменчивости «запад<=>восток». Корреляция с долготой составляет r=0.79. Карта делит генофонд на две зоны: западную и восточную. Эта карта чрезвычайно сходна с картой первой главной компоненты классических маркёров (рис. 8.2.1). Не только основные направления изменчивости, но размещение экстремумов компонент обоих типов признаков практически совпадает. Коэффициент корреляции между главными сценариями аутосомных маркёров - ДНК и классических - достигает r=0.81, подтверждая их удивительное сходство. Итак, первая компонента по ДНК данным выявляет ту же закономерность, что и по классическим маркёрам, поэтому естественно сохранить и ту же интерпретацию: паттерн ДНК полиморфизма в Восточной Европе сформировался главным образом в результате европеоидно-монголоидных взаимодействий. Основная черта обеих карт - выявление долготной изменчивости в восточноевропейском генофонде, подразделение его на западную и восточную зоны с постепенным переходом между ними. Различия карт касаются только особенностей этого перехода: если по ДНК данным значения компоненты изменяются довольно быстро, то по данным о классических маркёрах зона перехода занимает значительную территорию. Эти различия, думается, связаны с относительно небольшим числом популяций, изученных по ДНК маркёрам - в среднем 28 популяций, тогда как по классическим маркёрам - в среднем 103 популяции (табл.8.1.1.). Можно ожидать, что при увеличении числа популяций, изученных по ДНК маркёрам, зона перехода станет такой же плавной и широкой, как и по классическим маркёрам. ВТОРОЙ СЦЕНАРИЙ Вторая главная компонента ДНК полиморфизма (рис. 8.2.4) описывает 14.2% общей дисперсии и демонстрирует основное направление изменчивости «север<=>юг». Карта выявляет подразделение восточноевропейского генофонда на северную и южную зоны с плавным переходом между ними. Корреляция этой карты с географической широтой составляет r=0.70. Возможно, неслучайно это геногеографическое разделение соответствует физико-географическому разделению Восточной Европы на лесную и степную полосы. При этом в лесной зоне экстремум значений компоненты расположен на северо-востоке, в зоне расселения финно-угорских народов и северных русских популяций. Поскольку предполагается, что в формировании этих северных групп русских существенную роль сыграли финно-угорские племена (меря, чудь) [Алексеева, 1965, 1999], то можно предположить, что этот экстремум значений второй главной компоненты связан с зоной древнего расселения финно-угорских народов. Сравнивая эту карту второй компоненты по ДНК маркёрам с картой второй компоненты по классическим маркёрам (рис. 8.2.2), легко заметить их совпадение в основном направлении изменчивости. Коэффициент корреляции между вторыми сценариями аутосомных маркёров - ДНК и классических - достигает r=0.65, подтверждая их хорошее сходство. ТРЕТИЙ СЦЕНАРИЙ Карта третьей главной компоненты по ДНК маркёрам демонстрирует сложное распределение: четыре четких ядра (экстремумы значений третьей компоненты) легли по периферии, а в центре региона (в Волго-Окском междуречье, верхнем течении Дона и Днепра) взаимодействие этих ядер даёт промежуточные значения. Каждый из двух экстремумов компоненты занимает две разобщённые зоны, что и приводит к выявлению на карте четырех ядер. Таким образом, третьи сценарии и ДНК, и классических маркёров отражают сложность внутренней структуры восточноевропейского генофонда, что немаловажно после простых закономерностей первой и второй главных компонент. Итак, не только первые компоненты (коэффициент корреляции между ними r=0.81) и вторые компоненты (r=0.65) по ДНК и по классическим маркёрам, но и третьи компоненты оказываются сходными, выявляя одни и те же закономерности в генофонде: коэффициент корреляции между третьими компонентами составил r=0.73. Такая последовательность может вызывать только восхищение и уважение: оба типа маркёров генетики - вопреки всем капризам совершенно разной и неравномерной изученности - вторят друг другу даже во второстепенных и третьестепенных изгибах генетического ландшафта! Это важный аргумент в пользу необычайного сходства изменчивости ДНК и классических маркёров. СХОДСТВО СЦЕНАРИЕВ КЛАССИЧЕСКИХ И ДНК МАРКЕРОВ Подводя итог геногеографическому сравнению ДНК и классических маркёров, подчеркнём, что их сходство выявилось как статистическим анализом (совпадение уровней межпопуляционной изменчивости, раздел 8.1), так и картографическим анализом (высокие корреляции между картами главных компонент). Статистические сравнения этих двух типов маркёров проводятся во многих работах. А вот совпадение закономерностей в географической изменчивости ДНК и классических маркёров выявлено впервые. И обнаружено оно для всех трёх наиболее значимых главных компонент. Этот результат имеет и теоретическое значение. Он является новым важным подтверждением сходства межпопуляционной изменчивости ДНК и классических маркёров (см. Приложение). Сходство картин мира по ДНК и по классическим маркёрам удивительно! Причём совпадение карт главных сценариев служит более значимым аргументом, чем простой статистический расчёт. Ведь карты - намного более чуткий инструмент: они трехмерны (в отличие от одномерности статанализа), отражают более тонкие элементы структуры генофонда и пронизывают его на большую глубину. Одинаковость закономерностей по ДНК и по классическим маркёрам свидетельствует об объективности этих тенденций изменчивости восточноевропейского генофонда. Это также свидетельствует о целесообразности параллельного использования как ДНК данных, так и данных по классическим маркёрам. 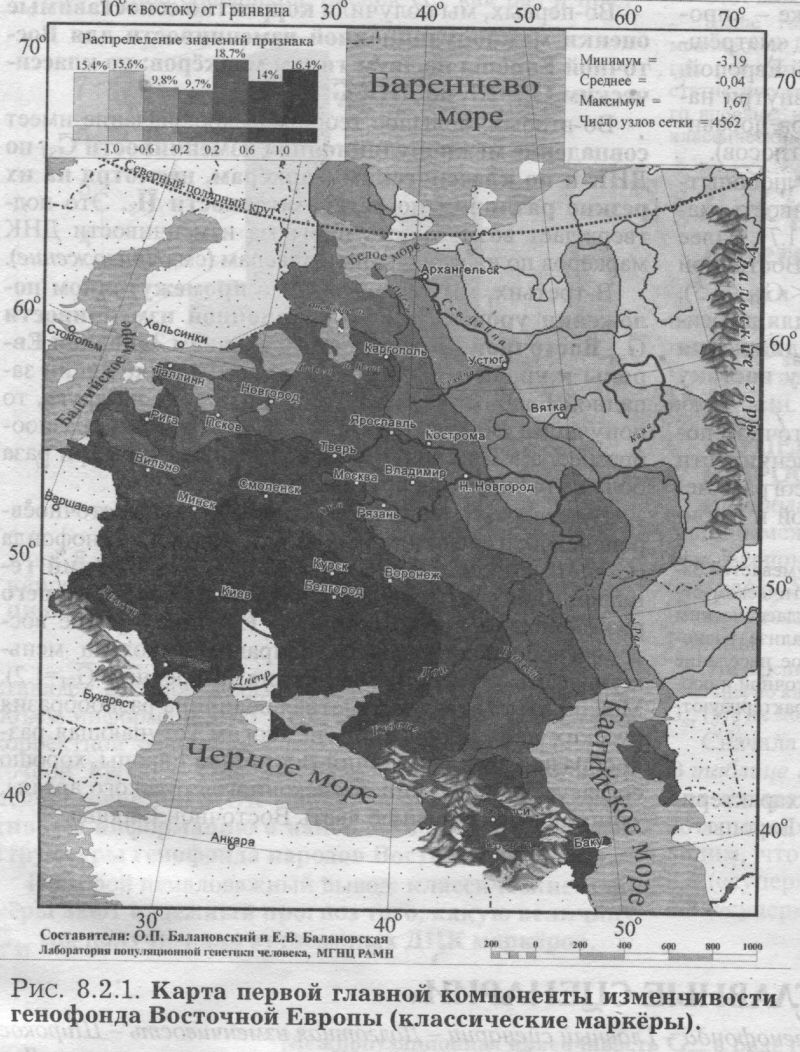
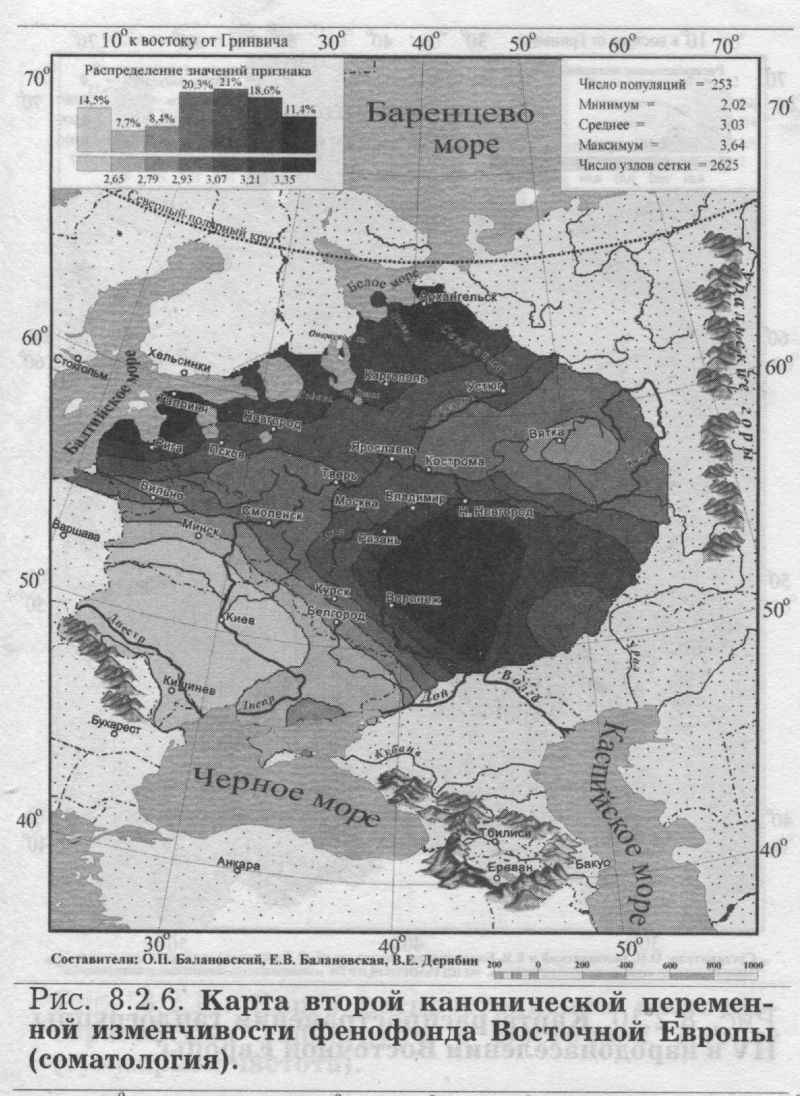
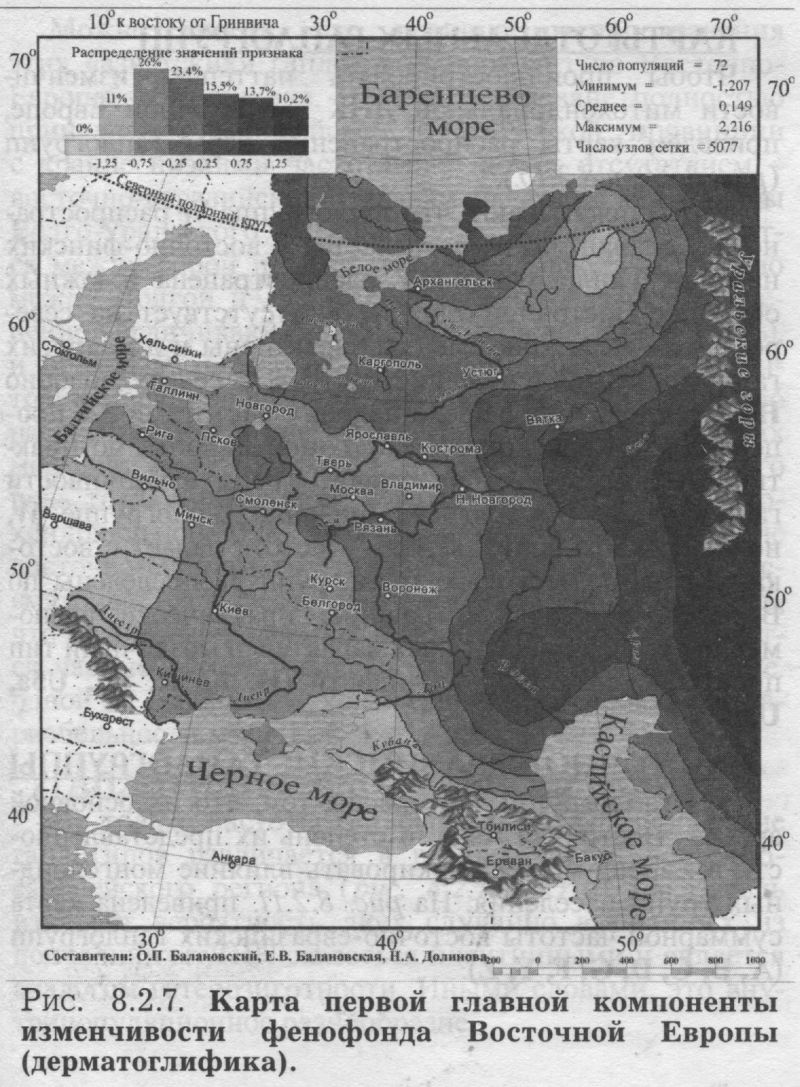
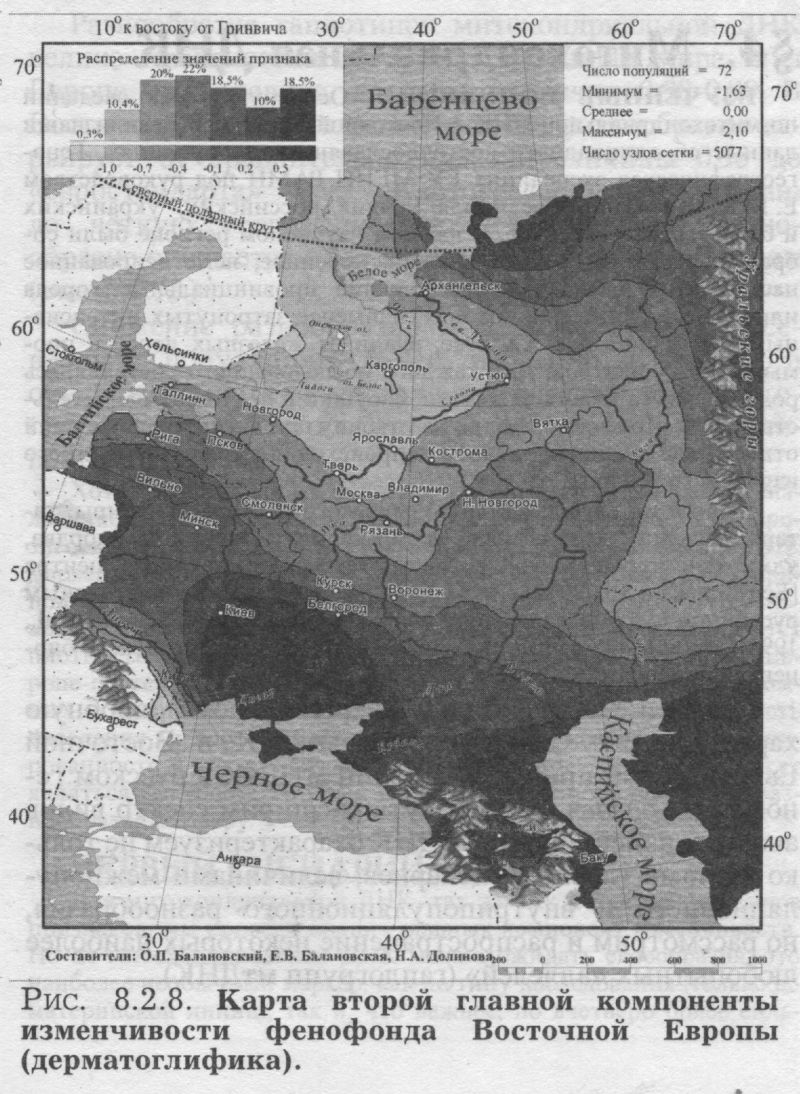
§3. Антропология: соматология и дерматоглификаПОЧЕМУ АНТРОПОЛОГИЯ Рассмотрев свидетельства двух очевидцев от генетики, мы можем перейти к свидетельствам антропологии. Её также представляют два очевидца - соматология и дерматоглифика (табл. 8.1.1). Какие же черты антропологии важны для геногеографа? 1) БОЛЬШОЙ НАБОР ПОПУЛЯЦИЙ. Антропологические признаки изучены по очень большому набору популяций. Если по ДНК маркёрам в Восточной Европе изучено в среднем около 30 популяций, по иммуно-биохимическим - примерно 100 популяций, то по данным соматологии - 253 популяции. По дерматоглифике изучено хотя и несколько меньше (72 популяции), но всё равно в два раза больше, чем по ДНК маркёрам. 2) ЕДИНЫЙ НАБОР ПРИЗНАКОВ. В отличие от генетики, все популяции изучены по одному и тому же набору антропологических признаков. А это очень важно! Это значительно повышает надёжность результирующих обобщённых карт. К тому же значения обобщённых признаков (главных компонент для дерматоглифики и их аналогов - канонических переменных для соматологии) рассчитывались по исходным, а не по интерполированным значениям: сначала стандартными статистическими методами были рассчитаны значения главных компонент, которые уже затем картографировались. Это исключает вопрос о ложных корреляциях (см. Приложение), который ставил под сомнение объективность расчёта главных компонент по картам, т. е. по интерполированным значениям [Sokal et al., 1999a,b; Rendine et al., 1999]. Забегая вперед, укажем, что сходство главных сценариев изменчивости генетики и антропологии вновь отвергает опасность ложных корреляций. 3) ЗРИМЫЙ ОБРАЗ ПОПУЛЯЦИЙ. За антропологическими признаками стоят не гены-невидимки, а зримый физический облик населения, детально разработанная антропологическая (расовая) классификация, наконец, все сообщения древних авторов и современные мнения о том, как выглядят представители тех или иных народов. Антропология всегда привлекается в исследованиях генофонда человека для объяснения обнаруженных фактов и закономерностей. Историческая антропология и в особенности дерматоглифика опираются на признаки с высокой наследуемостью. Это позволяет использовать эти признаки и в генетических исследованиях популяций. Поэтому, выявив направления изменчивости антропологических признаков, мы расширим знания о восточноевропейском генофонде. Более того, мы сможем более объективно интерпретировать изменчивость собственно генетических маркёров в терминах европеоидно-монголоидных пластов. АНАЛИЗ КАНОНИЧЕСКИХ ПЕРЕМЕННЫХ близок к методу главных компонент, оба они входят в одну группу статистических методов снижения размерности [Айвазян и др., 1989]. Основное различие между этими двумя методами состоит в том, что в каноническом анализе учитываются внутригрупповые корреляции признаков. Этот учёт внутригрупповых корреляций требуется только при использовании признаков соматологии, поскольку для дерматоглифики, как и для признаков генетики, внутригрупповые корреляции отсутствуют [Хить. 1983; Хить, Долинова, 1990]. Поэтому для анализа генетических и дерматоглифических признаков используют метод главных компонент [Cavalli-Sforza et al., 1994], тогда как для соматологических признаков обычно применяют канонические переменные [Дерябин, 2001], что и сделано в данной работе. Составление сводки по антропологическим признакам у народов Восточной Европы и канонический анализ проведены профессором кафедры антропологии МГУ В. Е. Дерябиным. Нами проведён картографический анализ этих результатов и прямое сопоставление закономерностей изменчивости антропологических и генетических маркёров. В отличие от других типов признаков, по данным соматологии сначала был проведён расчёт канонических переменных (аналогов главных компонент), а затем уже по этим данным создавались карты. ПЕРВЫЙ СЦЕНАРИЙ СОМАТОЛОГИИ На рис. 8.2.5. представлена география первой канонической переменной изменчивости признаков соматологии в коренном населении Восточной Европы. Карта первого сценария выявляет долготную изменчивость: один экстремум расположен на западе Русской равнины, другой - у народов Среднего и Южного Урала. Итак, по данным антропологии мы вновь видим тот же сценарий долготной изменчивости, что и по классическим, и по ДНК маркёрам. Для понимания смысла этой долготной изменчивости очень важно, что антропологические признаки, по данным о которых построена эта карта, лежат в основе расовой классификации. Поэтому теперь мы вполне уверенно и обоснованно можем интерпретировать долготную изменчивость в Восточной Европе в терминах взаимодействия европеоидной и монголоидной (уралоидной) рас. Мы только что рассматривали аналогичные карты долготной изменчивости по генетическим маркёрам, где мы также предложили «европеоидно-монголоидную» трактовку. Но доказательств, что дело именно в расовом составе населения, а не в другом факторе, изменяющемся с запада на восток, у нас не было. В случае же с антропологией «расовая» интерпретация изменчивости антропологических признаков естественно вытекает из того, что сама расовая классификация основана на этих признаках. И поскольку тот же долготный тренд выявлен по генетическим маркёрам, их изменчивость также следует объяснять тем же фактором. Привлечение данных антропологии позволило подтвердить правильность главного вывода, сделанного из анализа генетических данных. Коэффициент корреляции карт первой канонической переменной соматологии с картой первой главной компоненты по ДНК маркёрам высок (r=0.61), указывая на их сходство. Это сходство относится к совпадению долготного направления изменчивости. А размещение экстремумов, т. е. зон наибольшей «европеоидности» или «монголоидности», различно по антропологическим и генетическим данным. Но здесь уж лучше, видимо, положиться на антропологию - её карты достовернее, ибо они опираются на огромное число популяций, изученных каждая по всем признакам. ВТОРОЙ СЦЕНАРИЙ СОМАТОЛОГИИ Карта второй по значимости переменной (рис. 8.2.6.) выявляет четыре четких ядра с центральной областью их интенсивного взаимодействия. Подобную структуру мы уже встречали на карте третьей компоненты ДНК полиморфизма. Важным элементом рельефа таких карт является географическое положение «седловины» (промежуточных значений) - здесь эта центральная область расположилась на верхней Волге. ДЕРМАТОГЛИФИКА Мы видели, во-первых, принципиальное сходство в изменчивости различных типов признаков и, во-вторых, второстепенные, но весьма интересные различия между ними. Привлечение дополнительных данных - по иным типам признаков - не изменяя картину принципиально, позволяет уточнять многие факты, и, возможно, обнаруживать новые. В качестве такого дополнительного типа признаков использованы данные дерматоглифики. Дерматоглифические маркёры, в целом примыкая к другим антропологическим признакам, зачастую выявляют свои, особые закономерности в истории формирования народов [Хрисанфова, Перевозчиков, 1999]. Поэтому они широко используются во многих исследованиях популяций человека. Интерес генетиков к ним связан и с их высокой наследуемостью (её коэффициент варьирует от 0.6 до 0.8) - более высокой, чем для признаков соматологии [Хить, Долинова, 1990]. ПЕРВЫЙ СЦЕНАРИЙ ДЕРМАТОГЛИФИКИ Однако первая компонента даже столь специфических признаков (рис. 8.2.7.) демонстрирует ставшую уже привычной картину - долготный тренд, трактуемый как встреча европеоидного и монголоидного миров. ВТОРОЙ СЦЕНАРИЙ ДЕРМАТОГЛИФИКИ А вот карта второй компоненты дерматоглифических признаков своеобразна (рис. 8.2.8.). Рельеф этой карты выявляет различия между северо-западной областью - и всей остальной территорией Восточной Европы. Выявленное ядро, окружая Ладожское и Онежское озера, доходит до верхней Волги, Западной и Северной Двины. Подобная картина противопоставления одной области всем соседним обычно наблюдается на обобщённых картах другого типа - картах генетических расстояний, тогда как для карт главных компонент подобный тренд нетипичен. Действительно, среди многих опубликованных карт главных компонент [Cavalli-Sforza et al., 1994; Рычков, Балановская, 1990; Рычков и др., 1999; главы 4-9 данной книги] почти отсутствуют карты с таким типом изменчивости. Единственная несколько похожая карта главных компонент получена по данным о русских фамилиях [Балановский и др.. 20006]. География рассматриваемой второй главной компоненты (рис 8.2.8.) позволяет выдвинуть гипотезу - данные дерматоглифики обнаруживают своеобразие беломоро-балтийской расы, т. е. восточного варианта северо-европеоидной расы. Таким образом, включение в анализ ешё одного типа признаков (дерматоглифики) позволило не только подтвердить приоритет долготного тренда, но и выявить новую черту восточноевропейского генофонда - особость северо-запада, - которая не выявлялась в изменчивости иных признаков. 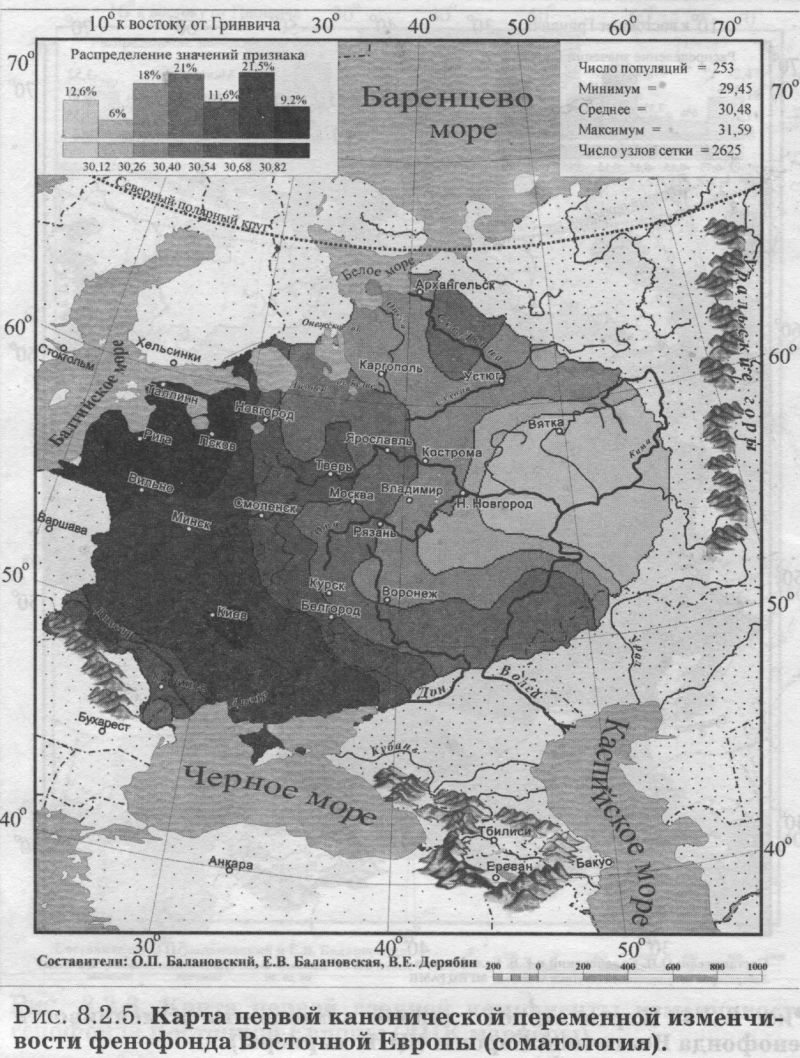
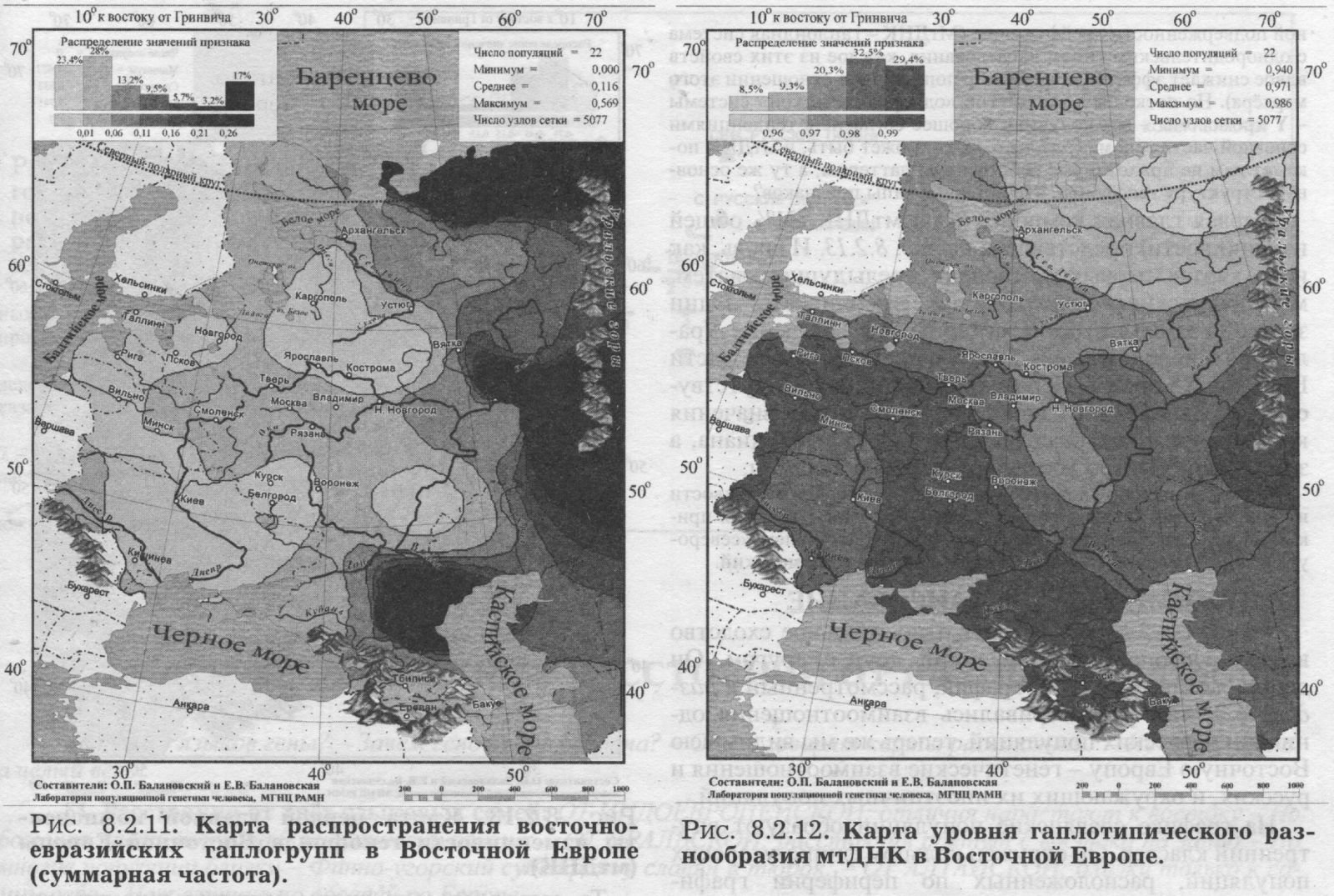
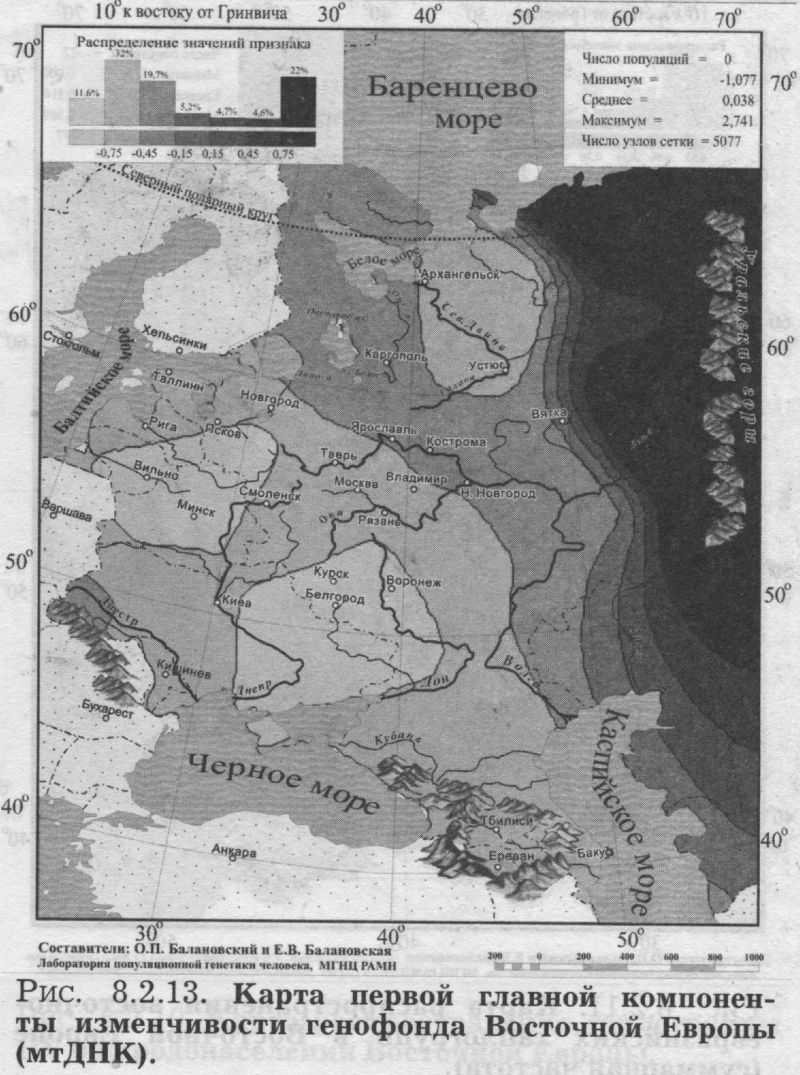
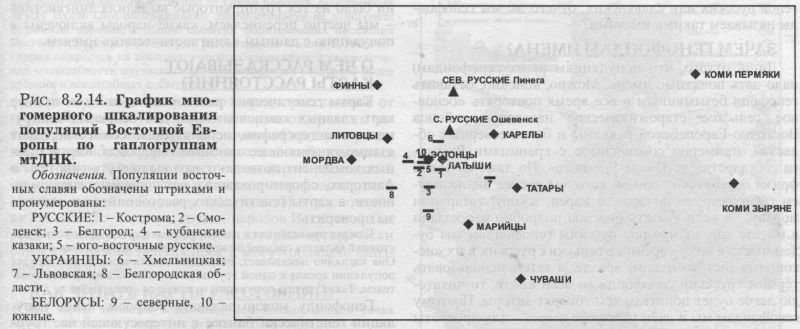
§4. Митохондриальная ДНК ИЗУЧЕННЫЕ ПОПУЛЯЦИИ. Основную часть сведений по митохондриальной ДНК в Восточной Европе составили наши данные об одиннадцати восточнославянских популяциях. Гено- географическая экспедиция ГУ МГНЦ РАМН под руководством Е. В. Балановской объединила усилия российских, украинских и белорусских генетиков. В каждом изучаемом регионе были собраны репрезентативные выборки: коренное, неметисированное население. Обследовались небольшие провинциальные города или сельское население в зонах, наименее затронутых интенсивными миграциями в XX веке, вдали от железных дорог и промышленных центров. Для каждого обследованного составлялась родословная (пробанд, родители, бабушки и дедушки), свидетельствующая, что все его предки на глубину не менее трёх поколений относились к данному этносу и происходили только из данного изучаемого региона. Данные по народам Волго-Уральского региона (башкиры, татары, чуваши, коми-зыряне, коми-пермяки, марийцы, мордва, удмурты) получены коллективом Уфимского научного центра [Бермишева и др., 2002]. Также использованы данные по северным русским популяциям [Bclyaeva et al., 2003], карелам [Sajantila et al., 1995]; эстонцам [Sajantila et al., 1995, 1996; данные Эстонского биоцентра] и ненцам [Saillard et al., 2000]. Полученные результаты впервые дают подробную характеристику изменчивости мтДНК в Восточной Европе. Как и при рассмотрении мтДНК в русском генофонде (раздел 6.2.) мы чуть расширим спектр видов анализа: изменчивость мтДНК охарактеризуем не только картами главных сценариев, величинами межпопуляционного и внутрипопуляционного разнообразия, но рассмотрим и распространение некоторых наиболее любопытных «аллелей» (гаплогрупп мтДНК). КАРТЫ ОТДЕЛЬНЫХ ГАПЛОГРУПП Чтобы проиллюстрировать паттерны изменчивости митохондриальной ДНК в Восточной Европе, приведём карты распространения пары гаплогрупп (рис. 8.2.9. и 8.2.10). Карты показывают, что гаплогруппа W распространена у славян, но не встречается у восточно-финских народов. Гаплогруппа HV распространена в южных областях Восточной Европы и отсутствует на севере. Столь же чёткие тренды характерны и для других гаплогрупп: U3 (снижение частоты к северу подобно HV); гаплогруппа I представлена с частотами 2-6 процентов в южных частях Восточной Европы, но практически отсутствует на севере. Тренд изменчивости гаплогруппы U4 долготен (подобно гаплогруппе W, но в обратном направлении - частота падает с востока на запад). Несколько гаплогрупп распределены по Восточно-Европейской равнине практически равномерно (Т, J), для некоторых характерен мозаичный тип пространственной изменчивости (Н, К, R, U2, U5a, U5b, V). ВОСТОЧНО-ЕВРАЗИЙСКИЕ ГАПЛОГРУППЫ Эти гаплогруппы интересны тем, что их основной ареал - Восточная Азия, и степень их представленности в Европе может маркировать влияние монголоидных групп населения. На рис. 8.2.11. приведена карта суммарной частоты восточно-евразийских гаплогрупп (А, В, С, D, G, F, Y, Z). Можно видеть, что граница распространения этих «азиатских» гаплогрупп проходит по ВосточноЕвропейской равнине. Восточные славяне полностью принадлежат европейской генетической провинции с крайне низкими частотами - почти отсутствием - восточно-евразийских гаплогрупп, тогда как народы Волго-Уральского региона существенно различаются между собой по частотам этих маркёров. Именно между Волгой и Уральским хребтом частоты постепенно увеличиваются до значимых величин (10 - 30%), и продолжают возрастать далее к востоку, на территории Западной Сибири. Бросается в глаза сходство рассмотренных выше карт главных компонент других маркёров (рис. 8.2.1., 8.2.3.. 8.2.5., 8.2.7.) и этой (своего рода тоже обобщённой, суммарной) карты для митохондриальной ДНК. Зная глобальное распространение восточно-евразийских гаплогрупп (оно подробно рассматривается в следующей главе 9) можно утверждать, что тренд, наблюдаемый в Восточной Европе, является частью общеевразийского долготного тренда, т. е. одной из основных закономерностей в глобальной вариабельности мтДНК. ГАПЛОТИПИЧЕСКОЕ РАЗНООБРАЗИЕ Не только частоты гаплогрупп, но и разнообразие гаплотипов различается в разных частях восточноевропейского региона (рис. 8.2.12). Этот показатель измеряет вероятность, двух случайно выбранных из популяции мтДНК оказаться идентичными и является аналогом гетерозиготности. Иными словами, это внутрипопуляционное разнообразие. Разнообразие гаплотипов митохондриальной ДНК велико практически во всех популяциях мира, и в Европе, как правило, достигает величин 0.98-0.99. В Восточной Европе гаплотипическое разнообразие варьирует от сравнительно низкой величины 0.95 до «евростандарта» 0.99. Картографирование выявило чёткую географическую изменчивость: разнообразие снижается от южных к северным популяциям (рис. 8.2.12.) Снижение разнообразия мтДНК от южных к северным популяциям ранее не было известно, и обнаружено нами благодаря последовательному картографированию всего комплекса показателей изменчивости мтДНК. Хотя в отношении мтДНК этот результат является новым, аналогичное снижение внутрипопуляционной изменчивости (гетерозиготности) к северо-востоку мы видели для классических и ДНК маркёров {рис. 8.1.1., 8.1.2.). Можно думать, что снижение внутри- популяционного разнообразия вызвано дрейфом генов, интенсивность которого, в свою очередь, в значительно степени задаётся плотностью населення. А плотность населения в Восточной Европе постепенно снижается в том же самом направлении: от южных к северным районам. Таким образом, сниженная плотность населения (и соответственно его большая дисперсность, изолированность и как следствие сильный генетический дрейф) могут являться фактором, который снижает внутрипопуляционное разнообразие к северу. ПЕРВЫЙ СЦЕНАРИЙ мтДНК Для митохондриальной ДНК, так же как и для прочих типов признаков, мы провели анализ главных компонент изменчивости. Но от митохондриальной ДНК можно ожидать своеобразия: это наиболее необычный маркёр, как по типу наследования (только по материнской линии), так и, что важнее, по вчетверо более сильной подверженности дрейфу генов. (МтДНК - гаплоидная система с однородительским типом наследования, каждое из этих свойств вдвое снижает эффективный размер популяции в отношении этого маркера). Но поскольку для другой «однородительской» системы - Y хромосомы - мы получили хорошее согласие с тенденциями основной части генома (раздел 6.3.), то, может быть, и мтДНК покажет нам не прихотливые собственные паттерны, а ту же основную структуру генофонда, что и прочие типы признаков? Первая главная компонента по мтДНК (31% обшей изменчивости) представлена на рис. 8.2.13. И вновь, как и на картах первых компонент предыдущих систем, мы обнаруживаем долготную изменчивость. Одни значения компоненты характерны для населения Урала, противоположные - для большей, западной части Восточной Европы. Переходная зона почти отсутствует - при движении с запада на восток низкие значения компоненты продолжаются вплоть до 50 меридиана, а затем очень быстро достигают высоких величин. Вторая компонента по мтДНК вбирает 17% общей изменчивости и демонстрирует более сложный рельеф. Значения задаются прикаспийским экстремумом, которому противопоставлен «североуральский» экстремум, и, в меньшей мере, причерноморский. МНОГОМЕРНОЕ ШКАЛИРОВАНИЕ Рисунок 8.2.14. показывает относительное сходство восточноевропейских популяций друг с другом. Он дополняет аналогичный график, рассмотренный в разделе 6.2: там рассматривались взаимоотношения одних лишь русских популяций, теперь же мы видим всю Восточную Европу - генетические взаимоотношения и русских, и окружающих их иноэтничных популяций. На графике славянские популяции образуют внутренний кластер. Он окружён кольцом финно-угорских популяций, расположенных по периферии графика. Кроме финноязычных групп, это внешнее кольцо включает также чувашей и северных русских; и именно в этих популяциях антропологи предполагают значительный финно-угорский субстрат. Расположение славян в центре финно-угорского кольца может отражать славянскую колонизацию нынешней Центральной России, сопровождавшейся ассимиляцией дославянского (финно-угорского) населения. МЕЖПОПУЛЯЦИОННАЯ ИЗМЕНЧИВОСТЬ Похожий результат получен и при анализе межпопуляционной изменчивости. В таблице 8.2.1. приведены средние генетические расстояния между популяциями, говорящими на языках основных лингвистических групп Европы. Для этого проанализированы данные по 67 европейским популяциям (информация нашего банка данных World Mitochondrial). Среди всех лингвистических групп Европы наиболее генетически разнообразны оказались финно-угры. За ними следуют романские и славянские народы. Германские народы - единообразны по митохондриальной ДНК. Таблица 8.2.1. 
То есть вновь мы видим значительную изменчивость в пределах финно-угорской общности, которая и на графике многомерного шкалирования образовала внешний кластер с большими расстояниям между популяциями. ИТОГИ Отметим основные черты восточноевропейского генофонда по данным о мтДНК: 1) Анализ изученных нами 11 восточнославянских популяций вместе с уже имеющимися в литературе данными по народам Волго-Уральского региона обнаружил географическую структурированность митохондриального генофонда в восточной половине Европы. Вновь, как и по другим типам признаков, основной закономерностью является долготная изменчивость. 2) В славянских популяциях встречены почти исключительно западно-евразийские гаплогруппы. Восточная граница зоны расселения славян является крайним западным рубежом для распространения «азиатских» гаплогрупп. 3) Среди популяций, представляющих основные языковые группы Европы (славянскую, романскую, германскую и финно-угорскую), наиболее дифференцированы финно-угры. 4) Данные по митохондриальной ДНК указывают на ассимиляцию восточными славянами митохондриального генофонда дославянского финно-угорского населения Восточной Европы, и особенно близкими к современным финно-угорским народам оказываются северные русские популяции. 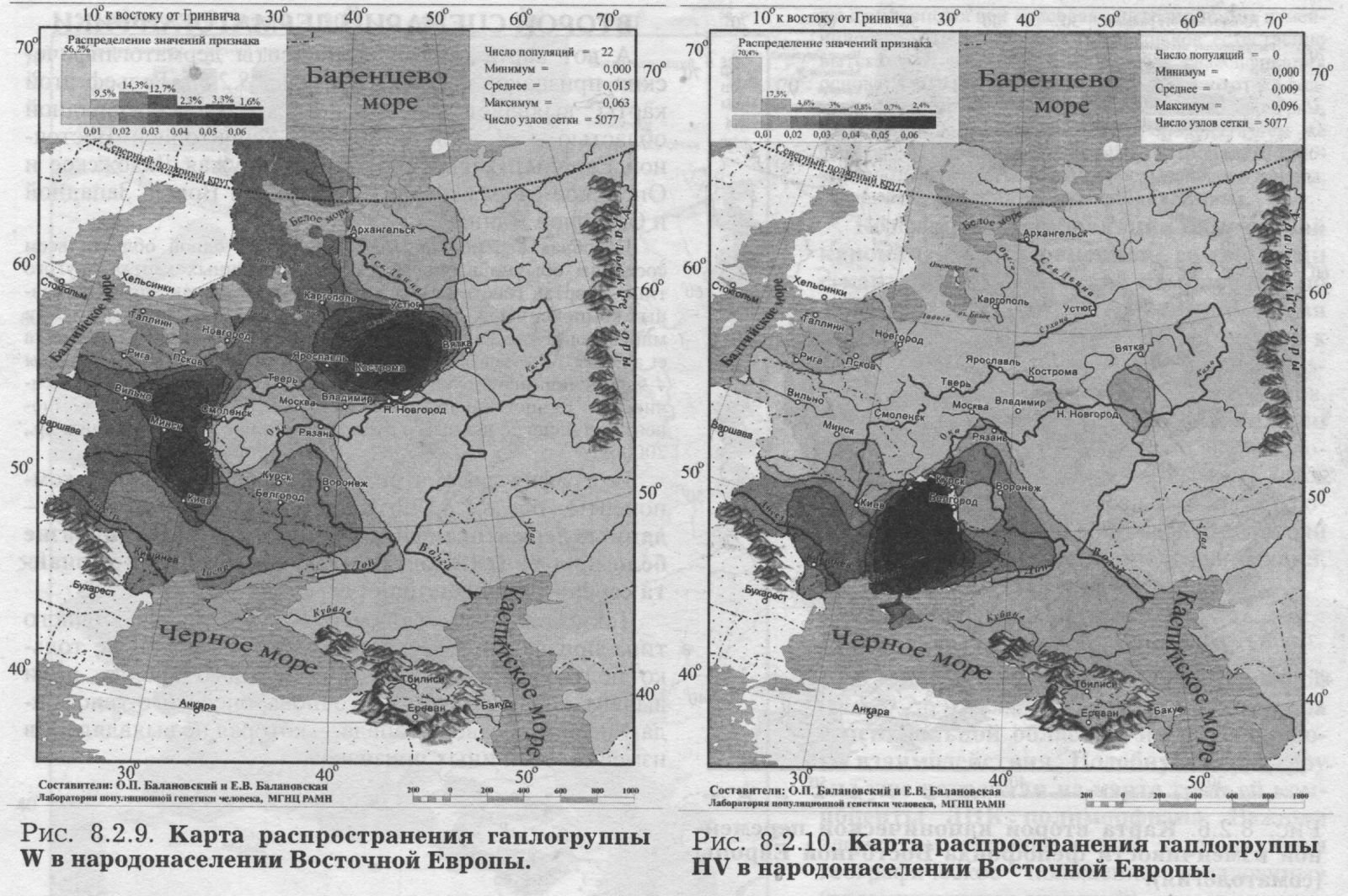
3При неэффективности метода главных компонент все компоненты описывают одинаковую часть общей дисперсии, то есть их информативность равна информативности исходных признаков. Если признаков 10. то каждый описывает десятую часть обшей дисперсии, если признаков сто - то сотую, и такая же дисперсия ожидается от главных компонент (новых признаков) при их неэффективности. В нашем случае хотя аллелей сто, но независимых признаков (число степеней свободы) меньше - 67, поскольку в каждом из 33 локусов частоты одного аллеля однозначно задаются частотами прочих аллелей (ведь сумма частот всех аллелей локуса всегда равняется единице). 100-33=67 независимых признаков, и дисперсия первой главной компоненты, ожидаемая при неэффективности метода составляет 1/67=0.149, то есть упомянутые полтора процента. 4Показатели ранговой корреляции Спирмена r приводятся по абсолютной величине, поскольку при корреляции с главными компонентами знак не имеет значения - для положительного и отрицательного полюса их знак условен, и они без последствий могут менять их на противоположные. |
загрузка...