6.3. Y хромосома (отцовские линии)
|
§1. Новейший маркёр в популяционной генетике: Романтический аргумент - Дробность выделения гаплогрупп - Часовая и минутная стрелки эволюции
§2. Изученные русские популяции: Фрагментарные литературные данные - И подробные собственные - Надежно изученный ареал - Полунаука §3. География гаплогрупп - взгляд извне и изнутри: Восемь главных гаплогрупп - «Славянская» R1a убывает к северу - «Северо-евразийская» N3 убывает к югу - «Балканская» I1b опять к северу - Сюрпризы викингов (I1a) - Странности европейцев (R1b) - Средиземноморские мозаики E3b, J2) - Уральская печать на русском генофонде (N2) §4. Гетерогенность русского генофонда по Y хромосоме: Выше, чем по мтДНК - Выше, чем по классическим маркёрам - Выше, чем у большинства народов Европы - Гетерогенность европейских народов по классическим и новейшим маркёрам §5. Русские популяции в географическом и в генетическом пространстве: Главный сценарий - Широтная изменчивость - От севера к югу - Север гетерогенен внутри себя Всем известно, как весома Для нас Игрек-хромосома: Хоть мала и некрасива, С нею вся мужская сила! М. Н. Монахова5 В самый разгар бума митохондриальных исследований, когда на изучении мтДНК сосредоточились усилия большинства популяционных генетиков, на сцену стремительно вышла другая генетическая система - Y хромосома. Хотя ей ещё не удалось потеснить мтДНК с позиций лидера, Y хромосома уверенно заняла место рядом с ней. Образовавшийся дуэт быстро стал принятым стандартом, и самым информативным сейчас считается одновременное изучение мтДНК и Y хромосомы, прочие маркёры считаются не столь важными. Отнюдь не умаляя значения прочих ДНК маркёров (некоторые типы которых, например аутосомные SNP гаплотипы, могут занять лидирующее положение уже в ближайшие годы) нужно признать, что сейчас львиная доля популяционных исследований приходится на мтДНК и Y хромосому. Полиморфизм Y хромосомы изучен уже в нескольких сотнях популяций по всему миру, уровень её изученности постепенно приближается к той же степени подробности, что и мтДНК. Ге- нотипирование Y хромосомы было стандартизовано в 2002, когда вышла статья с современной классификацией гаплогрупп [YCC, 2002]. Все нынешние исследования опираются на эту классификацию и постепенно уточняют её, в том числе путем внедрения новых маркёров. В этом разделе рассматриваются предварительные результаты, полученные авторами при изучении полиморфизма Y хромосомы в русском ареале. §1. Новейший маркёр в популяционной генетикеТы помнишь, что было в начале? Уже ты не помнишь, а жаль... Из двух половинок печали Сложилась большая печаль. Владимир Красногорский Формально, с методических позиций обе «половинки» - отцовская Y хромосома и материнская мтДНК - это простые системы нерекомбинирующих маркёров. Именно на явлении полного сцепления маркёров, входящих в эти системы, основаны новые методы, ставшие знаменем «новой популяционной генетики». Её своеобразными методами являются расчёт возраста гаплогруппы по разнообразию её субвариантов и возможность определять последовательность возникновения гаплогрупп, воссоздавать «родословное древо» гаплогрупп и гаплотипов. Других нерекомбинирующих систем (кроме мтДНК и Y хромосомы) в геноме человека нет; точнее, они лишь «частично нерекомбинирующие» и поэтому проблематичнее для использования. ДВЕ ПОЛОВИНКИ ПЕЧАЛИ Такая «методическая» аргументация в пользу внедрения Y хромосомы в арсенал генетических маркёров, несомненно, убеждает. Вместе с тем, в годы быстрого внедрения Y хромосомы имел хождение и «романтический» аргумент: если мтДНК, наследующаяся по женской линии, маркирует историю «женской части популяции», то Y хромосома, наследующаяся по мужской линии, есть её естественное дополнение - вторая половинка, рассказывающая об истории «мужской части популяции». И обе системы вместе опишут весь генофонд популяции. И сейчас можно встретить веру в то, что митохондриальная ДНК описывает одну половинку генофонда, Y хромосома - другую, и из двух этих половинок можно сложить целый генофонд. Этот романтизм оставил след в неуклюжей терминологии вроде «мужского генофонда» (Y хромосома присутствует только у мужчин). Однако, с точки зрения классической популяционной генетики, тендерное подразделение генофонда («по полу») выглядит спорным и грустным. Ведь мы изучаем не отдельные гены и не отдельных людей, а популяции человека, которые не могут быть «мужскими» или «женскими». При этом широко известно, что обе системы - мтДНК и Y хромосомы - могут не «дополнять» друг друга, а смещать оценки параметров генофонда в одну и ту же сторону. Это ясно видно хотя бы из того, что обе системы являются гаплоидными и «однородительскими». Поэтому дрейф генов действует на них в четыре раза сильнее, чем на аутосомные маркёры. Соответственно изменяются и оценки ряда генетических параметров популяции - в первую очередь столь важного, как оценка гетерогенности генофонда. Это приводит к тому, что генофонд через призму мтДНК может искажаться примерно тем же образом, что и через призму Y хромосомы. И тогда совместное использование этих маркёров может послужить не их взаимной проверке, не взаимодополнению, а взаимному подтверждению искажённых (по сравнению с основной частью генофонда) оценок. Из этого уже «сложится большая печаль». Видимо, самая безопасная стратегия - пользоваться самыми разными маркёрами и обдуманно компенсировать недостатки одних параллельным использованием других маркёров, лишенных этих конкретных недостатков. Это вселяет надежду сохранить объективность и независимость своих выводов. ИЗУЧЕНИЕ Y ХРОМОСОМЫ Популяционная изученность Y хромосомы уже сейчас приближается к уровню мтДНК. Но необходимо учитывать, что объёмы выборок по Y хромосоме, как правило, существенно меньше, поскольку изучается ДНК лишь мужчин. А они, если не прилагать специальных усилий, составляют обычно менее половины выборки. К тому же результаты первых работ (примерно до 2000 года) сейчас уже малопригодны, поскольку набор используемых для генотипирования маркёров был слишком невелик. Так что сейчас Y хромосоме следует отвести второе место. Но потенциально её разрешающая способность (возможность выделять все более дробные гаплогруппы) намного выше, чем мтДНК, и это привлекает к Y хромосоме все большее внимание исследователей. Генотипирование Y хромосомы состоит в определении, к какой гаплогруппе относится каждый образец ДНК. Доказательством принадлежности к гаплогруппе служит наличие специфического для данной гаплогруппы маркёра (или маркёров). Как правило, это SNP маркёры - замена одного из нуклеотидов Y хромосомы. Поскольку классификация гаплогрупп представляет собой иерархическое, ветвящееся дерево, то становится особо важным, с каким разрешением проведено генотипирование: определена ли принадлежность образца к гаплогруппе, или к её определенному варианту, или же с точностью до субварианта и так далее. SNP и STR - ЧАСОВАЯ И МИНУТНАЯ СТРЕЛКИ ЭВОЛЮЦИИ Определение гаплогрупп Y хромосомы проводят по набору диаллелъных маркёров (обычно SNP) - точно так же, как гаплогруппы мтДНК определяются диаллельными ПДРФ маркёрами в её кодирующей части («часовая стрелка»). Что же касается гипервариабельного сегмента мтДНК («минутная стрелка»), то и у Y хромосомы есть его аналог — набор микросателлитных локусов (STR). Подобно тому, как ГВС1 в мтДНК состоит из быстро (по сравнению с ПДРФ-маркёрами) мутирующих сайтов, так и микросаттелиты Y хромосомы мутируют намного быстрее SNP-маркёров: они соотносятся как минутная и часовая стрелки эволюции. И если в мтДНК каждая гаплогруппа, определенная по ПДРФ-маркёрам, вмещает в себя целый веер линий, выявленных по ГВС1, то и каждую SNP-гаплогруппу Y хромосомы можно подразделить на множество гаплотипов, определяемых по панели микросаттелитов. Ноприэтомдля мтДНК разрыв между ГВС1 и ПДРФ-маркёрами не так велик, как аналогичный разрыв между двумя типами маркёров Y хромосомы. Для мтДНК, хотя и не со стопроцентной вероятностью, но все же возможно определить гаплогруппу по мотиву в ГВС1. Для Y хромосомы корреляция между микросателллитным гаплотипом и SNP-гаплогруппой меньше, хотя её удаётся выявить [Rootsi et al., 2004]. В большинстве работ изучаются только SNP маркёры и соответственно распределение гаплогрупп Y хромосомы. Что же до микросаттелитных маркеров, то их анализируют реже - по минутной стрелке не определишь, который час. Ими пользуются для оценки возраста в пределах отдельных гаплогрупп и некоторых других видов анализа [Roewer et al., 2005]. §2. Изученные русские популяцииЛИТЕРАТУРНЫЕ ДАННЫЕ Как и большинство генетических маркёров, Y хромосома наиболее изучена в Европе. В том числе есть данные и по русским популяциям (табл. 6.3.1.). В работе [Wells et al., 2001] представлены северные русские (N=49; русские Архангельска, проживающие там не менее трёх поколений [Евсеева, личное сообщение]), поморы (N=28; студенты Архангельска, все предки которых происходят из поморских сел [Евсеева, личное сообщение]) и русские Ташкента (N=89). В статьях [Rosser et al., 2000] и [Karafet et al., 2002] представлены данные по русским «вообще» без указаний о географической привязке популяций (объёмы выборок N=122 и N=61. соответственно). Ни в этих статьях, где русские были лишь одним из многих изученных этносов, ни в других известных нам работах не предпринимались попытки изучить русский генофонд по маркёрам Y хромосомы6. Надо заметить, что и самих этих данных, как по их суммарному объему (N=349), так и особенно по географической приуроченности (фактически описаны лишь «русские-неизвестно-откуда» и «северные русские») для выявления структуры генофонда явно недостаточно. НАШИ ДАННЫЕ Независимо от перечисленных работ, мы целенаправленно провели анализ полиморфизма Y хромосомы в русских популяциях, изучив 1228 индивидов из 14 популяций (табл. 6.3.1.). Наше экспедиционное обследование было спланировано так, чтобы охватить коренное русское население из многих географически различных регионов, но не выходя за пределы «исконного» ареала русского народа. Все дальнейшее изложение опирается только на полученные нами данные. Проблема «исконного» русского ареала рассматривалась в главах 4 и 5. Основными участниками и/или организаторами экспедиций, проведенных под общим руководством Е. В. Балановской в 2000-2005 гг., были О. П. Балановский, И. В. Евсеева, Э. А. Почешхова, М.И. Чурносов. Также участвовали М. Н. Болдырева, О. Бобрецова (северные популяции), И. Н. Лепендина, М. С. Жерлицына, И. Н. Сорокина. В. Ю. Песик, Н. А. Рудых (южные популяции), А. С. Пшеничное (западные популяции). Генотипирование маркёров Y хромосомы выполнено О. П. Балановским при участии С. Роотси на базе Эстонского биоцентра, возглавляемого Р. Виллемсом. Определены частоты гаплогрупп Y хромосомы на основе типирования SNP-маркёров. Информация из опубликованных источников не используется, как говорилось, из-за неясной географии популяций, а для северно-русских выборок - из-за их небольших объёмов. Надо сказать, что 14 популяций - не так мало даже в сравнении с хорошо изученными классическими маркёрами (в среднем 35 русских популяций, но карты включались в анализ, если изученных популяций было хотя бы 10) или дерматоглификой (28 популяций). Благодаря наличию этих данных, Y хромосома становится одним из наиболее изученных ДНК маркёров для русских популяций. Для сравнения - по мтДНК надёжно изучено в два раза меньше популяций (7 популяций в «исконном» ареале). Полученные данные уникальны - ни для одного другого народа мира пока нет столь обширных данных. На примере русского народа впервые можно увидеть, каковы могут быть различия «отцовских» генов между разными популяциями одного и того же народа. ГЕОГРАФИЯ ИЗУЧЕННЫХ ПОПУЛЯЦИИ География изученных русских популяций обширна (табл. 6.3.1). Еще лучше, чем из таблицы, это видно на карте. Четырнадцать изученных популяций отмечены темными кружками на всех картах (рис. 6.3.1.-6.3.9.). Возникает вопрос, весь ли «исконный» русский ареал мы изучили, обследовав эти популяции? Ответ можно получить, воспользовавшись методом карт надёжности, подробно описанном в Приложении. Такая карта надёжности для наших 14 популяций представлена на рис. 6.3.1. Эта карта показывает, какие территории могут считаться охваченными генетическим изучением. Тс территории. которые расположены далеко от изученных популяций, представлены как зоны недостоверного прогноза - по нашим данным мы не можем надёжно прогнозировать частоты гаплогрупп на этих территориях. Например, среди изученных популяций нет ни одной из Волго-Окского междуречья. Но наличие данных по окружающим регионам (Тверская, Костромская, Орловская области) делает интерполяцию на междуречье достаточно надежной. Однако на северо-западе (Ленинградская и запад Архангельской областей), как показывает карта надёжности, данных не достаёт: для этой территории мы не можем дать надёжного прогноза. НАДЁЖНЫЙ АРЕАЛ Картографирование частот гаплогрупп (карты на рис. 6.3.2.-6.3.9.) проведено лишь для территории, надёжно обеспеченной исходными данными (надёжная область на рис. 6.3.1). Поэтому северо-западные части «исконного» русского ареала остались в некартируемой области. Напротив, на юге в картируемую область вошла популяция кубанских казаков, которая, строго говоря, находится за пределами «исконного» ареала. За этими исключениями, область, в которой картографирована изменчивость Y хромосомы, примерно соответствует ареалу, в пределах которого анализировались и антропологические признаки, и классические маркёры, и фамилии (карты в главах 4, 5, 7). ВЫДУМКИ ЖУРНАЛИСТОВ К сожалению, эта карта ешё до выхода нашей книги появилась в журнале «КоммерсантЪ Власть», где была истолкована в прямо противоположном смысле: будто бы чисто техническая карта надёжности, просто заданная расположением наших изученных популяций, определяет ареал русского народа [КоммерсантЪ Власть, 2005; №38]. Это было бы смешно, если бы мы могли посмеяться вместе с читателями. Нам же пришлось на это и другие столь явно ошибочные утверждения давать опровержения [см. на сайте www.genofond.ru]. Ведь так очевидно, что границы русского ареала могут очертить только гуманитарные науки, а генетика может лишь послушно планировать свои исследования в очерченных ими географических рамках. Но уже не смешно, а опасно, когда простому планированию полевых исследований, по произволу журналистов, придаётся сила научного свидетельства об историческом ареале народа. Это и есть «...полунаука, самый страшный бич человечества, хуже мора, голода и войны, не известный до нынешнего столетия. Полунаука - это деспот, каких ещё не приходило до сих пор никогда. Деспот, имеющий своих жрецов и рабов, деспот, пред которым все склонилось с любовью и суеверием, до сих пор немыслимым, пред которым трепещет даже сама наука и постыдно потакает ему» [Ф. М. Достоевский. «Бесы»]. Поэтому, чтобы не потакать геополитическим спекуляциям, нам пришлось «обрезать» надёжно изученный ареал русского народа по современной политической границе России (рис. 6.3.1). 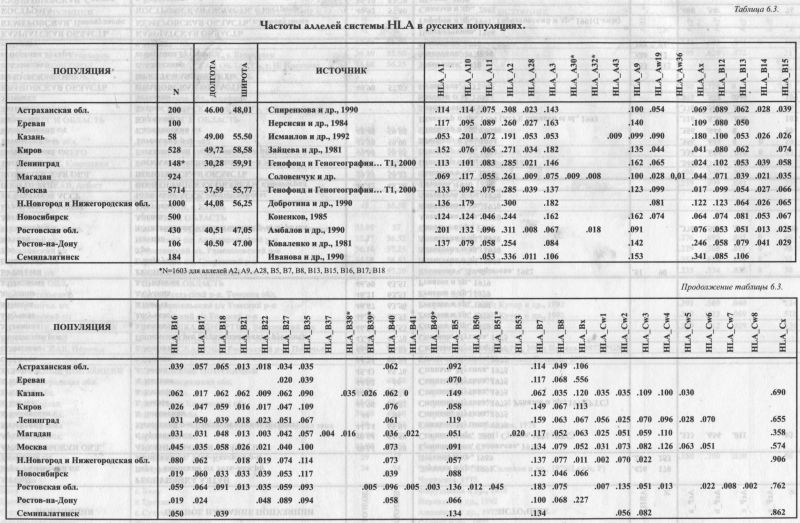

§3. География гаплогрупп: взгляд извне и изнутри В этом параграфе мы осмотрим генетический портрет русского народа на фоне Европы. Знание того, как гаплогруппы Y хромосомы распространены в глобальном масштабе, помогает понять, что означает их присутствие (или отсутствие) у русских. Это взгляд на русский генофонд извне. А различия в частотах гаплогрупп между самими русскими популяциями выявляют внутреннюю структуру русского генофонда - это мы назовём взглядом изнутри. Оба взгляда мы будем бросать на одни и те же карты (рис. 6.3.2.-6.3.9). Сначала на карте мы будем рассматривать общеевропейскую закономерность в распространении гаплогруппы, а потом её изменчивость внутри «надёжного» русского ареала (оконтурен серой линией на всех картах). Исходные данные для «европейского» картографирования описаны в Приложении (раздел 5). Известно, что гаплогруппы Υ хромосомы географически специфичны, причём в намного большей степени, чем аутосомные маркёры и мтДНК. Многие гаплогруппы Υ хромосомы имеют конкретные, чёткие ареалы. Если гаплогруппы мтДНК называют «континент-специфичными», то гаплогруппы Υ хромосомы часто приурочены к более дробным географическим регионам. Поэтому именно Υ хромосома многообещающа для изучения изменчивости внутри этноса, для выявления различий между близкими популяциями, которые неотличимы по другим маркёрам. Многие гаплогруппы, обнаруженные в коренных русских популяциях, редки и не удовлетворяют 1% критерию полиморфизма: лишь восемь гаплогрупп встречаются со средней частотой выше 1% (табл. 6.3.2). Но сумма частот этих восьми гапдогрупп составляет 95%, что даёт нам право во всех видах анализа учитывать лишь эти гаплогруппы, опуская прочие редкие варианты. Рассмотрим географию этих восьми гаплогрупп. 
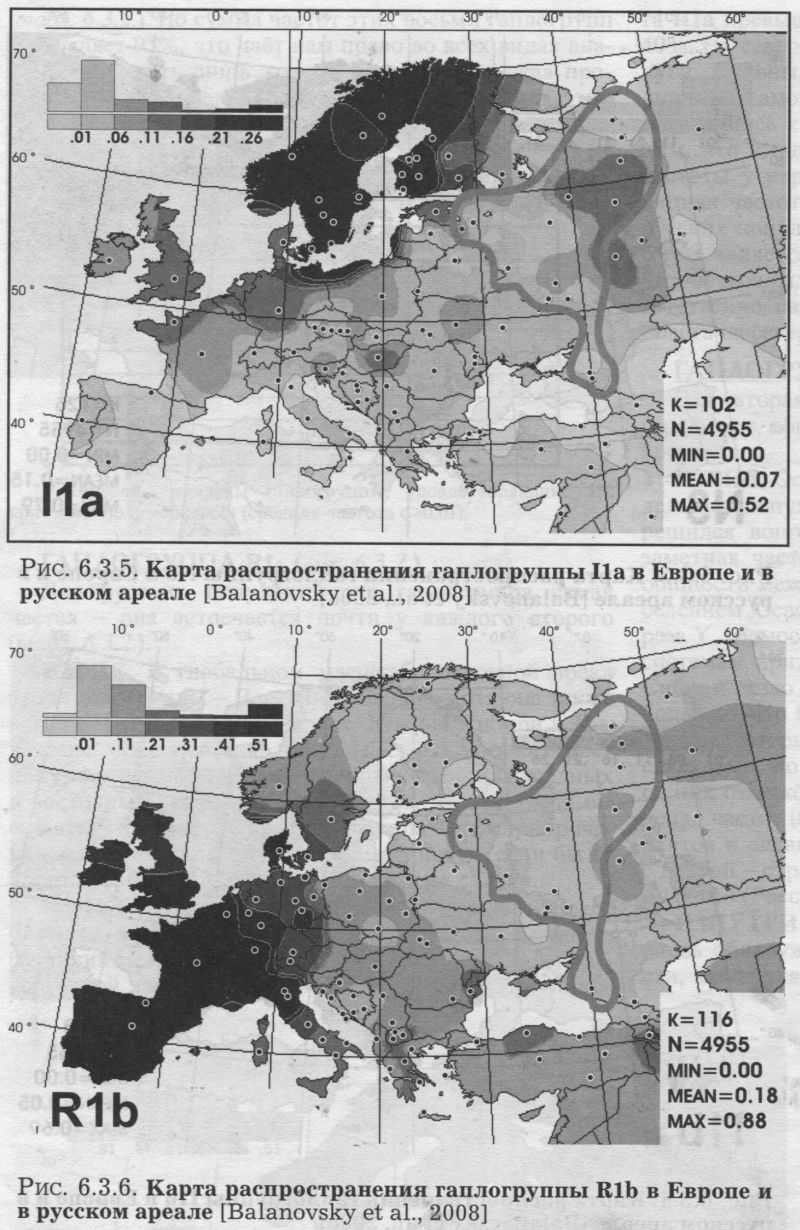
ГАПЛОГРУППА R1a (рис. 6.3.2.) Для русских популяций эта гаплогруппа самая частая - она встречается почти у каждого второго (табл. 6.3.2.). ИЗВНЕ. В глобальном масштабе основной ареал этой гаплогруппы - Восточная Европа, хотя она также встречается в Западной Европе, Южной Сибири и Индостане. «Европейская» карта (рис. 6.3.2.) показывает приуроченность этой гаплогруппы к ареалам западных и восточных славян, а также балтов. Можно было бы сказать, что в Европе эта гаплогруппа чётко приурочена к балто-славянской языковой общности, если бы её частота не была резко снижена у южных славян. ИЗНУТРИ. В пределах изученного русского ареала (область, оконтуренная на карте серой линией) видны различия между большей западной и меньшей северовосточной частями генофонда. На западе и юге частота Rla превышает 50% и лишь местами снижается до 40%. На северо-востоке (за Волгой и в бассейнах Ветлуги, Сухоны, Вычегды) частота Rla ниже 40%. И лишь на самом севере (на Мезени) вновь возрастает. Вернувшись к западной зоне высоких частот, мы увидим, что с юга её ограничивает некоторое снижение частоты у украинцев. При этом в белорусских популяциях частота Rla столь же велика, как в западных русских популяциях. Пока остаётся открытым вопрос о том, является ли гаплогруппа Rla маркёром расселения восточных славян, а её сниженная частота, соответственно, свидетельством мощного финно-угорского пласта на северо-востоке русского ареала. ГАПЛОГРУППА N3 (рис. 6.3.3.) Эта вторая по частоте гаплогруппа в русском генофонде, вбирает в себя пятую часть (19%) фонда Υ хромосом. ИЗВНЕ. Основной ареал N3 - Сибирь (до 90% в отдельных популяциях) и Восточная Европа. Как бы ни решился вопрос о прародине этой гаплогруппы, её заметная частота у русских показывает некую долю общности между русским генофондом и коренным населением Северной Азии. Такая общность по маркёрам Υ хромосомы неожиданна, поскольку по другим системам признаков генофонды Восточной Европы и Сибири резко различны (см., например, главу 9). Любопытно, что балтоязычные популяции (латыши и литовцы), которые по Rla решительно объединялись со славянами, по гаплогруппе N3 ещё более решительно от них отмежёвываются. Карта показывает резкий перепад частот (более чем на 20%!) примерно по границе балтов и славян: с одной стороны Латвии и Литвы, и с другой - стран с преобладанием славяноязычных популяций (России, Белоруссии и Польши). ИЗНУТРИ. В пределах русского ареала для N3 вновь обнаруживается картина широтной изменчивости, как и для Rla. Мы опять видим на карте две области. Только их значения обратные предыдущей карте: высокие частоты на севере, низкие - на юге. Удивителен довольно резкий перепад в частотах N3 между севером и югом и строго широтная граница между зонами высоких и низких частот. Данные по смежным народам подтверждают, что N3 приурочена к северной половине Восточной Европы. Русский генофонд тем самым полностью подчиняется восточноевропейской закономерности о концентрации N3 к северу от 57 параллели. И снова, учитывая редкость N3 у белорусов и украинцев, можно обсуждать толщину финно-угорского пласта в русском генофонде и его залегание преимущественно на севере. ГАПЛОГРУППЫ I1а и I1b (рис. 6.3.4. и 6.3.5.) Эти две сестринские гаплогруппы можно назвать, ориентируясь на зоны их максимальных частот, Скандинавской и Балканской гаплогруппами. Их распространение в русском ареале может помочь нам обнаружить связи русского населения с этими двумя регионами. ИЗВНЕ. Частота I1а максимальна в Скандинавии, и постепенно снижается к югу. Напротив, I1b распространена главным образом на Балканах, и её частота постепенно снижается к северу. Вся центральная полоса Европы оказывается зоной интерференции этих гаплогрупп. У русских преобладает «балканская» гаплогруппа, хотя частота «нордического» варианта также существенна. Соотношение I1a:I1b составляет 1:2. При этом у белорусов и украинцев «нордическая» гаплогруппа, по сравнению с «балканской», практически незаметна. Соотношение I1a:I1b составляет 1:5 для украинцев и 1:15 для белорусов [Пшеничное, Атраментова, Ищук, Чурносов, Тегако, Виллемс, Балановская, неопубл. данные]. Для украинцев естественна близость с географически соседним населением Балкан. ИЗНУТРИ. Внутри русского ареала распространение «Балканской» гаплогруппы I1b (рис. 6.3.4.) закономерно: частота плавно падаёт с юга (>13%) на север (0%). То есть, тренд lib снова широтный. Но только изменение частоты lib не резкое, как для N3, а постепенное. Мы видим, что тренд, обнаруженный в русском генофонде, полностью встраивается в более общий, панъевропейский тренд распространения гаплогруппы lib. Но её сестринская гаплогруппа уже не подчиняется европейской закономерности. Генетический рельеф «Скандинавской» гаплогруппы I1a (рис. 6.3.5.) в русском ареале оказывается неожиданным. Казалось бы, высокие значения 11а должны быть вблизи Скандинавии, на северо- западе русского ареала. Там, а также по западной границе можно ожидать «варяжское» влияние в виде повышенных частот I1a. Однако компактный локальный максимум I1a (11-12%) вырисовывается совсем в другой области, на северо-востоке. Это ядро ясно выделяется на фоне окружающих низких частот (менее 6%), свойственных всему остальному русскому ареалу. Реальность этого ядра повышенных значений опирается на данные по трём русским популяциям, изученным по обширным выборкам. Конечно, по сравнению с частотами в Скандинавии (25-40%) этот локальный максимум второстепенен. Но его удаленность от основной зоны высоких частот гаплогруппы в Скандинавии требует объяснений. Трудно предположить тесные связи Скандинавии именно с Заволжьем и бассейном Вычегды, помимо прочих русских территорий. Видимо, популяционная история гаплогруппы I1a сложнее, чем простая экспансия из Скандинавии, и, возможно, включает древние связи между финно-угорскими племенами Восточной Европы и предками германоязычных скандинавов. ГАПЛОГРУППА R1b (рис. 6.3.6.) ИЗВНЕ. R1b свойственна населению Западной Европы, что ярко выявляется на карте. Присутствие гаплогруппы у русских выражено в той же небольшой степени, что и в целом для популяций Восточной Европы (например, 6% у украинцев и 3% у белорусов). ИЗНУТРИ. В русском ареале Rib встречается с частотой от нуля до 13%, в среднем составляя 6%. В отличие от предыдущих гаплогрупп, в её распределении по русскому ареалу чёткой географической закономерности нет. Для большинства популяций характерны средние частоты. Повышенные частоты отмечены в трёх популяциях на востоке, пониженные - в одной на северо-востоке. Учитывая, что основная зона R1b - Западная Европа, неожиданно, что её частота в русском ареале возрастает скорее к востоку, чем к западу. Впрочем, эта слабая тенденция может быть случайной. Тем не менее, она подчёркивает отсутствие заметных влияний на русский генофонд из Западной Европы, которые должны были бы выразиться на карте в клинальной изменчивости частоты «западноевропейской» гаплогруппы. ГАПЛОГРУППА Е3b (рис. 6.3.7.) ИЗВНЕ. Е3b характерна для Северной Африки (до 80%) и Средиземноморья в его самом широком понимании. В Европе наибольшие частоты отмечены на Балканах (10-20%) [Semino et al., 2004]. До русского ареала доходит лишь «затуханье сахарской волны» (Н. Гумилев). ИЗНУТРИ. Внутри русского ареала генетический ландшафт состоит в повышении частоты в центральной полосе и снижении к северу и к югу (подобные ландшафты мы уже не раз встречали среди самых разных признаков - генетики, фамилий, антропологии). Такой паттерн лишь отчасти согласуется с европейским трендом для этой гаплогруппы (снижение к северу). ГАПЛОГРУППА J2 Соис. 6.3.8.) ИЗВНЕ. J2 преобладает на Ближнем Востоке и на Кавказе [Wells et al., 2001; Semino et al., 2000]. В русском генофонде она могла появиться не только со стороны Кавказа, но и Южной Европы (включая Балканы), где J2 также нередка [Semino et al., 2004]. В Европейском масштабе J2 следует тому же генетическом рельефу, что ЕЗЬ: максимум в Средиземноморье и снижение к северу. ИЗНУТРИ. Но «русский» ландшафт гаплогруппы J2 (рис. 6.3.8.) оказывается гомогенным в пятипроцентной шкале карты. Когда карта была перестроена в более дробной шкале, выявился ряд ядер повышенных частот на востоке, средние частоты на юге и отсутствие гаплогруппы на севере. Но средний объём наших выборок приближается к 100 образцам на популяцию. При таких выборках доверительный интервал для частоты 2% приближается к наблюдаемому размаху её изменчивости. Это лишает основательности долгие рассуждения, поскольку тренды становятся неотличимы от случайных шероховатостей рельефа. Для анализа географии таких редких признаков нужно особое везение: наличие очень чёткого тренда, очень большие выборки или обилие изученных популяций. ГАПЛОГРУППА N2 (рис. 6.3.9.) ИЗНУТРИ. Гаплогруппа N2 как раз характеризуется очень чётким трендом. Хотя её средняя частота столь же невелика, как для ЕЗЬ и J2, её распространение географически чётко приурочено. Гаплогруппа N2 встречена только на Русском Севере, в трёх популяциях Архангельской области. ИЗВНЕ. Учитывая, что .основной ареал N2 - угорские народы Западной Сибири, а в Европе она встречается только среди волго-финских народов, финно-угорский отпечаток на генофонде Русского Севера виден по N2 рельефнее, чем по любому другому маркёру. ИТОГИ ИЗВНЕ. Итак, по спектру гаплогрупп Y хромосомы и их средним частотам русский генофонд укладывается в более крупный регион Восточной Европы (доминирование восточноевропейских гаплогрупп Rla и N3). Видны связи практически со всеми смежными регионами: Западной Европой (наличие Rib), Скандинавией (11а), Балканами (lib, J2, ЕЗЬ), Западной Сибирью/Уралом (N2). Лишь связь с Кавказом выявляется слабо, поскольку гаплогруппа G (наиболее частая у северокавказских народов), практически не встречается у русских и лишь чуть более часта у украинцев. ИЗНУТРИ. Однако при таком видении русский генофонд выступает лишь точкой, которую мы сравниваем с другими точками - соседними регионами. Поэтому мы рассматривали не только средние показатели, но и карты распространения этих гаплогрупп в русском ареале. Это позволило увидеть внутреннюю гетерогенность русской «точки», занимающей большую часть Восточной Европы с десятками миллионов обитателей. По трём самым частым гаплогруппам (Rla, N3, lib), а также для редкой N2 в русском ареале обнаружилась широтная изменчивость, согласующаяся с общеевропейскими трендами этих гаплогрупп. Распространение в русском ареале умеренно частых гаплогрупп Rib и На носит сложный характер и не согласуется с европейскими закономерностями. Наконец, редкие для русских гаплогруппы J2 и ЕЗb распределены в русском ареале мозаично. 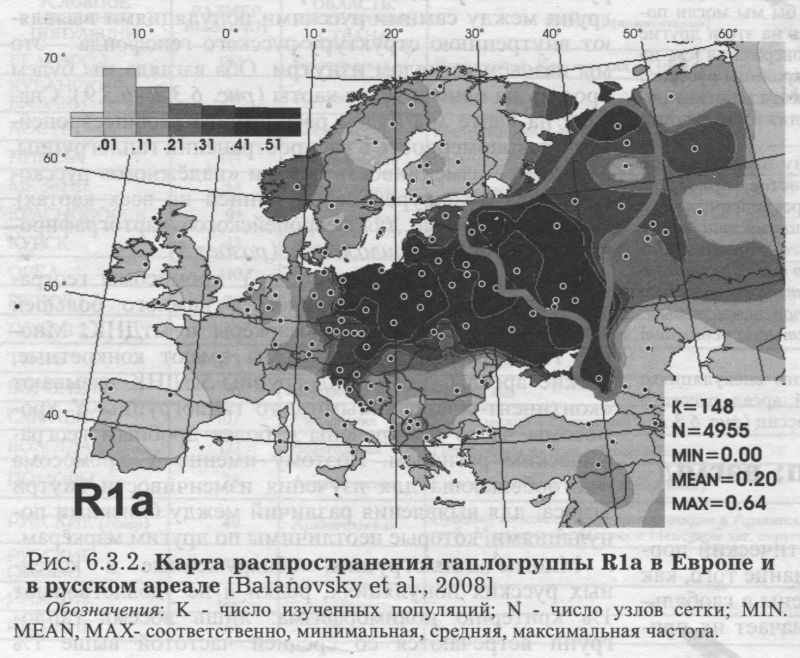


§4. Гетерогенность русского генофонда по Y хромосомеТРИ МЕТОДА ОБОБЩЁННОГО АНАЛИЗА В предыдущем разделе мы воспользовались уникальным свойством Y хромосомы - высокой географической специфичностью её гаплогрупп, их приуроченностью к конкретным территориям. Это позволило выявить связи разных частей русского генофонда с соседними группами народонаселения. Однако каждая из гаплогрупп рисует свой эскиз портрета генофонда. видит только одну из множества его проекций, отражается в одной из граней кристалла. Чтобы выявить наиболее общие черты портрета генофонда, присутствующие подспудно в каждом эскизе, надо перейти к обобщённому анализу изменчивости Y хромосомы. Для этого мы применим обычный арсенал методов, который широко использовался и в предыдущих главах. Сначала (§4) количественно определим гетерогенность русского генофонда - величину межпопуляционных различий. И так же, как мы это делали для классических маркёров (глава 5), оценим гетерогенность по шкале «много» - «мало». Затем в §5 рассмотрим карты главных компонент изменчивости по маркёрам Y хромосомы точно так же, как по признакам антропологии (глава 4) и классическим маркёрам (глава 5). И в заключение, оценим взаимное расположение русских популяций на графике многомерного шкалирования - так, как мы уже рассмотрели аналогичный график для мтДНК. ГЕТЕРОГЕННОСТЬ Y ХРОМОСОМЫ. мтДНК И АУТОСОМНЫХ МАРКЕРОВ Чтобы оценить размах межпопуляционных различий по Y хромосоме, для неё так же, как и для мтДНК (см. 6.2. §3), была рассчитана матрица генетических расстояний М. Нея между 14 русскими популяциями. Ни для одного другого народа мира ещё не проводилось количественное измерение внутриэтнической изменчивости Y хромосомы в сравнении с изменчивостью других народов. Средние генетические расстояния по гаплогруппам Y хромосомы составили d=0.142. Этот показатель оценивает гетерогенность генофонда, его внутриэтническую изменчивость. Различия между русскими популяциями по частотам гаплогрупп Y хромосомы (d=0.142) оказались в четыре раза больше, чем по мтДНК (d=0.033, см .раздел 6.2.). ПАТРИЛОКАЛЬНОСТЬ. Это ожидаемый результат, поскольку во многих работах (на уровне межэтнических сравнений) показано, что изменчивость Y хромосомы намного более выражена, чем мтДНК [Seielstad et al., 1998; Rosser et al., 2000; Semino et al., 2000]. Это явление нередко связывают с патрилокальностью браков (т. е. переездом жены на родину мужа - при этом отцовские гены остаются на родине отца), тогда как при матрилокальности изменчивость Y хромосомы (отцовские гены «мигрируют» на родину матери), напротив, становится меньше, чем по мтДНК [Oota et al., 2001; Hamilton et al., 2005; Destro-Bisol et al., 2004]. Пaтрилокальность, характерная для большинства народов, приводит к тому, что гены, наследующиеся по мужской линии, из поколения в поколение остаются в том же месте, тогда как мтДНК благодаря брачным миграциям интенсивно перемещается вместе с женщинами с их родины на родину мужа. Рассмотрим теперь гетерогенность русского народа по аутосомным маркёрам (классическим). При расчёте тем же методом средних генетических расстояний она составила d=0.013. Используя строго одну и ту же меру гетерогенности, можно смело утверждать, что различия между русскими популяциями по Y хромосоме (d=0.142) намного выше (рис. 6.3.10.), чем по мтДНК (d=0.033) и по классическим маркёрам (d=0.013). Можно рассудить, что если различия выявляются легче, то Y хромосома - удобная система для изучения структуры генофонда. 
Примечание: Средние генетические расстояния между всеми русскими популяциями по трём типам маркёров. СРАВНЕНИЕ С ДРУГИМИ НАРОДАМИ Итак, изменчивость русских популяций по Y хромосоме на порядок выше, чем по аутосомным маркёрам. Но надо ещё ответить на вопрос: велика или мала гетерогенность русского народа по Y хромосоме в сравнении с другими народами? Полученная оценка гетерогенности (d=0.142) характеризует л/елспопуляционные, но вжтирыэтнические различия: различия между русскими популяциями (то есть между популяциями внутри этноса). Ряд исследований показал, что изменчивость по Y хромосоме действительно велика, но почти все эти работы описывают только межэтническую изменчивость - различия между народами, а не внутри их. Поэтому мы провели анализ внутриэтнической изменчивости в Европе (табл. 6.3.3.), обобщив опубликованные данные. В анализ включены только народы, изученные по четырём и более популяциям с объёмом выборок более 70 человек в каждой популяции. Чтобы удовлетворить этому условию, популяции с небольшими объёмами выборок объединялись с соседями. (Для русских были объединены популяции «Пинега» и «Мезень», «Псков 1» и «Псков 2», «Тверь» и «Кострома», «Курск» и «Орел».) Гетерогенность русского генофонда занимает третье место в ряду одиннадцати изученных народов Европы (табл. 6.3.3.). Конечно, «места», занимаемые генофондами по степени их гетерогенности, могут измениться по мере изучения других народов Европы (и более подробного изучения тех же самых народов). Но уже сейчас можно сделать общий вывод, что по данным об Y хромосоме русский генофонд оказывается одним из наиболее гетерогенных этнических генофондов в Европе. ГЕТЕРОГЕННОСТЬ НАРОДОВ ЕВРОПЫ: ДВА ТИПА МАРКЕРОВ Таблица 6.3.3., суммирующая данные о гетерогенности по Y хромосоме, аналогична таблице 5.3.1., где гетерогенность рассчитана по классическим маркёрам. Для сопоставимости этих таблиц мы рассчитали гетерогенность Y хромосомы не только как средние расстояния, но и через близкородственную статистику - более «помехоустойчивую» величину GST Сравнение этих таблиц позволяет сделать три вывода: 1. Изученность Y хромосомы на внутриэтническом уровне пока оставляет желать лучшего - накоплены данные лишь о 11 народах, изученных в среднем по шести популяциям, а по классическим маркёрам проанализированы данные о 31 народе Европы, и эти народы изучены в среднем по 14 популяциям. 2. Гетерогенность народов по Y хромосоме оказывается почти в четыре раза выше, чем по классическим маркёрам. В среднем по Европе внутриэтническая изменчивость Y хромосомы составляет 3.3 (как следует из данных таблицы 6.3.3.), тогда так по классическим маркёрам она составляет 0.9 (расчёт по данным таблицы 5.3.I., для сопоставимости из этой таблицы мы учитываем только европейские народы). 3. Несмотря на разницу в абсолютных величинах внутриэтнической изменчивости, её относительная величина у разных народов похожа по обоим типам маркёров. Например, гетерогенность русских и итальянцев высока и по Y хромосоме, и по классическим маркёрам, а гетерогенность поляков занимает одно из последних мест как по Y хромосоме (GST=1.1), так и по классическим маркёрам (GST=0.3). Отметим также, что по Y хромосоме (табл. 6.3.3.) изменчивость популяций в пределах одного народа, хотя и велика, но значительно (в пять раз!) меньше, чем изменчивость среднеэтнических частот гаплогрупп между рассмотренными народами Европы. Это означает, что «дифференцирующая мощность» гаплогрупп Y хромосомы с увеличением географических дистанций только растет! ЧТО ЛУЧШЕ - ГЕНЕТИЧЕСКИЕ РАССТОЯНИЯ ИЛИ GST? Гетерогенность каждого народа мы оценили двумя мерами: как средние генетические расстояния между каждой парой популяций и как межпопуляционная изменчивость GST. Хотя по абсолютной величине генетические расстояния в среднем оказываются в три раза выше, чем Gsr но относительные величины обеих мер хорошо согласуются друг с другом. Кажущееся расхождение в случае турок вызвано необычно высоким уровнем филогенетического разрешения, с которым изучен этот народ (генотипирование проводилось на 26 гаплогрупп, тогда как для остальных народов - только на 8-12 гаплогрупп). Если же привести данные по туркам к общему уровню (8 гаплогрупп), то средние генетические расстояний уменьшаются с 16.5 до 3.5, а величина GST оказывается более устойчивой к изменению уровня филогенетического разрешения, сокращаясь с 1.3. только до 0.8. Более привычные молекулярным генетикам показатели AMOVA очень близки к GST и мы их здесь не приводим. 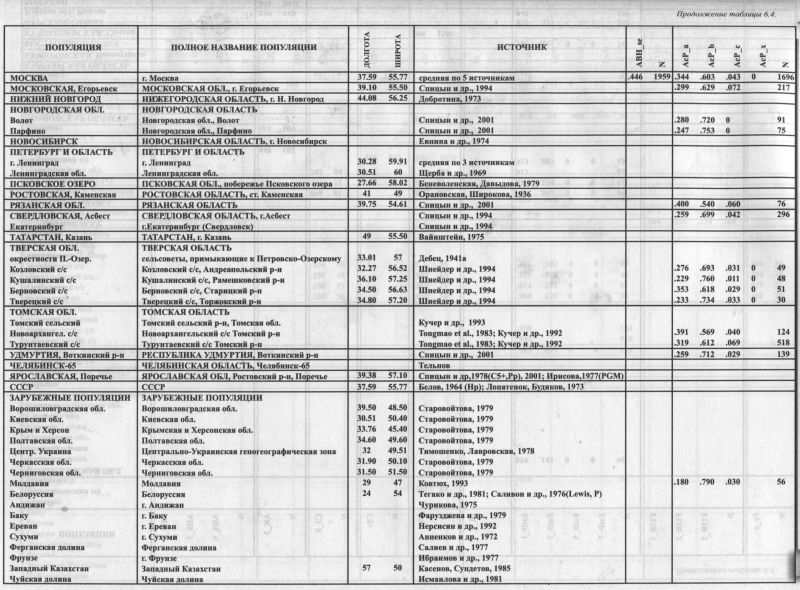
§5. Русские популяции в географическом и в генетическом пространстве Мы определили величину гетерогенности русского генофонда по маркёрам Y хромосомы. Теперь рассмотрим, как эта межпопуляционная изменчивость реализована в географическом пространстве (карты главных компонент) и в генетическом пространстве (график многомерного шкалирования). РУССКИЕ ПОПУЛЯЦИИ в ГЕОГРАФИЧЕСКОМ ПРОСТРАНСТВЕ Карты главных компонент изменчивости Y хромосомы рассчитаны по тем же восьми картам, которые рассматривались в предыдущих разделах, и обобщают изменчивость этих восьми гаплогрупп. Карты главных компонент построены по той же технологии, что и карты главных компонент по классическим маркерам, дерматоглифике и фамилиям (главы 5. 4, 7). Конечно, объём данных (число популяций и число гаплогрупп) по Y хромосоме меньше, чем по этим системам признаков, но всё же достаточен для проведения компонентного анализа. ПЕРВЫЙ СЦЕНАРИЙ. Карта первой главной компоненты (рис. 6.3.11) выявляет ландшафт широтной изменчивости русского генофонда. Эта картина уже знакома нам по данным соматологии, дерматоглифики и классическим генным маркёрам. Значения компоненты плавно изменяются от южных русских популяций к северным. Это означает, что таково основное направление генетических различий по маркёрам Y хромосомы - компонента описывает 40% общей изменчивости. Первая главная компонента высоко коррелирует с шестью из восьми гаплогрупп (табл. 6.3.4). Это говорит о том, что широтная изменчивость - действительно «повсеместный» тренд для системы Y хромосомы. Сходство этого тренда с аналогичными трендами по другим системам признаков очевидно (рис. 4.4.3., 4.4.4., 5.3.2). Полиморфизм Y хромосомы в русских популяциях подчиняется той же господствующей тенденции широтной изменчивости, что и прочие системы маркёров. Надо отметить, что преобладание широтной изменчивости выявляется именно обобщённой картой. На картах отдельных гаплогрупп четкая широтная картина видна лишь дважды (для Rla картина не полностью широтная), а на остальных картах столь явно не проявляется. Использование компонентного анализа позволяет сделать этот завуалированный тренд очевидным. ВТОРОЙ СЦЕНАРИЙ. Второе по значимости направление изменчивости рисует довольно сложную картину: три ядра положительных значений на юге, северо-западе и севере, и ядро отрицательных значений на востоке. Подобные «четырёхядерные» ландшафты характерны для третьих компонент генофонда Восточной Европы (см. главу 8). Карту мы не приводим, так как важность этой компоненты невелика. Она описывает четвертую часть (26%) общей изменчивости всех восьми гаплогрупп, то есть её значимость лишь в два раза выше каждой из исходных карт отдельных гаплогрупп. РУССКИЕ ПОПУЛЯЦИИ В ГЕНЕТИЧЕСКОМ ПРОСТРАНСТВЕ График многомерного шкалирования (рис. 6.3.12.) показывает относительное сходство всех 14 популяций друг с другом по частотам гаплогрупп Y хромосомы. (Этот график построен точно так же, как и график по гаплогруппам мтДНК, рассмотренный в §3 раздела 6.2.). В нижней части графика виден плотный кластер из восьми популяций, тогда как остальные шесть популяций разбросаны далеко друг от друга в верхней части графика. Важно то, что все популяции, образовавшие компактный кластер, происходят из южной и центральной части русского ареала. И наоборот, все северные популяции генетически удалены друг от друга, занимая разные углы верхней половины графика. Характерно, что в нижней половине графика (рис. 6.3.12.) оказались те популяции, которые на карте первой главной компоненты (рис. 6.3.11.) входят в зону положительных значений, а популяции, которые на карте обладают отрицательными значениями компоненты, на графике отошли в его верхнюю часть. На графике своеобразен северо-запад - одна из псковских популяций примыкает к южному кластеру, тогда как вторая вошла в «северную» зону. Основная закономерность, видимая на графике, состоит в том, что на юге сходство популяций намного больше, чем на севере ареала. ИТОГИ Из такого чёткого соответствия между географией популяций и генетическими расстояниями между ними можно сделать два вывода. Во-первых, мы вновь убедились в значимости различий между севером и югом русского ареала. Во-вторых, на севере различия между популяциями намного выше. Центральные и южные русские популяции в сравнении с Русским Севером кажутся близнецами. 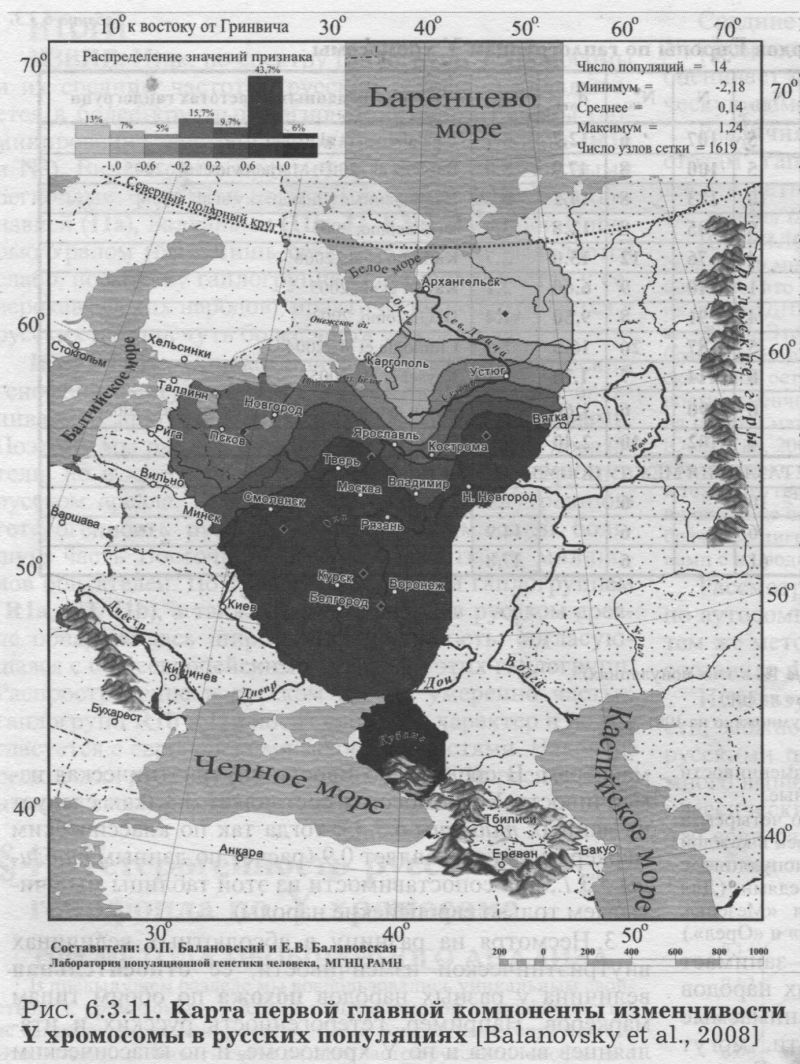
5 Публикуется впервые, с любезного разрешения автора. 6 Пока эта глава готовилась к печати, вышла статья Б. А. Малярчука с данными по пяти русским популяциям (табл. 6.2.2.). |
загрузка...