5.2. «Простые» карты классических маркёров
|
Источники информации и маршрут - Прогулка по «матрёшкам» - Карта - Сравнение с соседями - Гипотезы отбора и истории
§1. Биохимические маркёры. HP: Широтная изменчивость - GC: Широтная на глобусе, но не на русской карте - PI: Долготная в Евразии и в русском генофонде (Р1*М2) - Аномалии вне «исконного» ареала - TF: Разный отбор на разные аллели? - Мозаика в Европе - Широтные волны в русском ареале (TF*C2) - ACPI: АСР1*А возрастает к юго-востоку - Но частоты у юго-восточных соседей снижены! - PGM1: Долготная изменчивость в Евразии - Мозаика в русском ареале - ESD: Широтные волны - Незаметно влияние соседей - GLOl: Убывает к периферии русского ареала - Ноу соседей обратные тренды! - 6PGD: Сниженная изменчивость §2. Группы крови. MN: «Матрёшки» мира - «Матрёшки» Северной Евразии - Гомогенность Восточной Европы - Снижение у русских Мезени - RH*d: Евразийская карта — Долготная из.менчивость в Европе - Мозаика в русском ареале - За деревьями исчезает лес - АВО: Евразийские карты - «А» убывает на восток - «В» убывает на запад - «О» убывает к середине — Русские карты - «А» убывает на северо-восток - Лоскутное одеяло «В» - «О» убывает на юго-запад - Русский Север - Противоречия по MN и АВО - Глобальная закономерность не обязывает этносы к её исполнению - Этнос в своем ареале может её отменить! §3. Риск несовместимости матери и плода по группам крови: Несовместимость по Резус-фактору - Несовместимость по АВО - Учет обеих систем - Сто пятнадцать промежуточных карт - Итоговая карта риска - Отличия генетического риска от реальной заболеваемости §4. Обзор «простых» карт: Четкие закономерности для нескольких генов - Сложная география многих генов - Или вовсе мозаика - Окраинные аномалии необязательно связаны с соседями - Принцип «матрёшки» Напомним ещё раз, что география отдельных генов - не самоцель нашей книги. Карты отдельных генов, рассматриваемые в этом разделе, нужны нам лишь для того, чтобы представлять, сколь велико реальное разнообразие генетических рельефов, свернувшееся потом в генетический ландшафт обобщённой карты генофонда. Но и без «простых» карт разных генов обойтись никак нельзя. Без внимательного рассмотрения карт отдельных аллелей представление о пространственной изменчивости генофонда неизбежно страдает излишней абстрактностью, а порой приводит к переоценке универсальности выявленных закономерностей. ИСТОЧНИКИ ИНФОРМАЦИИ В этом разделе мы детально рассмотрим изменчивость каждого из классических маркёров в русских популяциях. Источником информации послужил банк данных «Русский генофонд» (Приложение, раздел 5). Частоты аллелей и оценки межпопуляционных различий GST для популяций русского народа, для пяти регионов Северной Евразии и для девяти регионов мира приведены в Приложении (раздел 6) и на сайте www.genofond.ru. Но главным источником информации для нас служат не сами частоты аллелей в русских популяциях, а основанные на них карты (рис. 5.2.1.-5.2.22.). Карты выявят географию каждого из маркёров, покажут, каковы закономерности его изменчивости в русском ареале. К таблицам же, содержащим частоты генов в русских популяциях, мы будем изредка обращаться лишь затем, чтобы уточнить, что представляет собой популяция, заинтересовавшая нас на карте. Например, если популяция резко отличается от соседей, то стоит проверить, не вызвано ли такое отличие просто малым объёмом её выборки? Другим источником информации служат результаты статистического анализа карт. Они сведены в таблице 5.2.1. К ним относится, во-первых, GST - степень изменчивости частоты аллеля, рассчитанная по карте. Эту степень изменчивости данного маркёра мы будем сравнивать со средним уровнем изменчивости, рассчитанным по картам всех классических маркёров (GST = 1.36). Во-вторых, статистический анализ карт даёт нам частные корреляции частоты аллеля с долготой, широтой и множественный коэффициент корреляции с географическими координатами (табл. 5.2.1). Они рассчитаны по специальным корреляционным картам (см. Приложение), а в таблице указаны среднее, минимальное и максимальное значения коэффициентов корреляции на такой карте. В-третьих, приведены и основные статистические показатели: минимальная, средняя и максимальная частоты аллеля, встречающиеся на картах. Величина дисперсии не приводится, поскольку приведён близкий по смыслу показатель GST. Наконец, есть и третий источник информации, которым мы будем пользоваться. Это частоты классических маркёров у соседних народов или в соседних регионах. Источником этих сведений служит банк данных «GENEPOOL» (Приложение, раздел 5), в разделе 6 Приложения приведены средние частоты генов и разнообразие GST для пяти регионов Северной Евразии5. Таким образом, мы приводим практически всю исходную информацию и результаты анализа: исходные данные (Приложение, раздел 6) и результаты анализа на сайте и в табл. 5.2.1., карты на рис. 5.2.1.-5.2.22. Поэтому читатель находится в равных условиях с авторами, располагая той же информацией. И все описания изменчивости классических маркёров, то есть всё последующее изложение в этом разделе 5.2., читатель может проверить, переосмыслить... или переписать иначе. 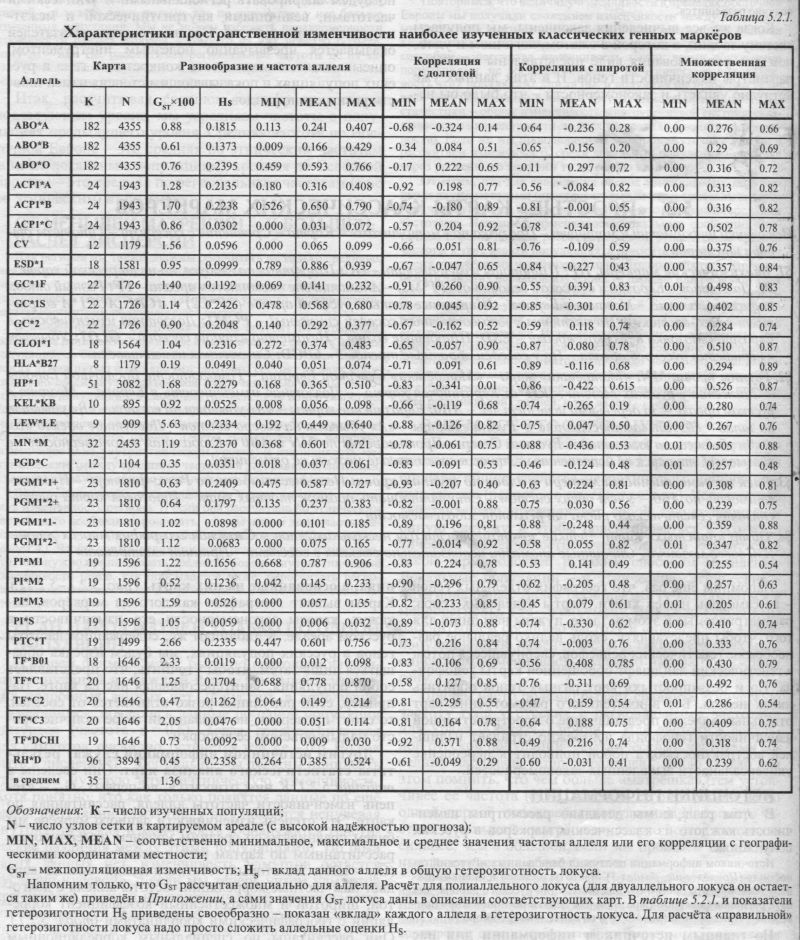
«МАРШРУТ» РАССМОТРЕНИЯ КАРТ В описании каждой карты мы старались следовать определенному плану. 1) СПРАВКА. В начале каждого подраздела приводится очень краткая справка о данном гене: его функция, географическое распространение в глобальном масштабе. Эта информация взята главным образом из монографии [Генофонд и геногеография...], при подготовке которой в 1986-1994 годах (с деятельным участием одного из авторов), было учтено и компилировано всё множество доступных на тот момент источников информации по классическим маркёрам. 2) ИЗМЕНЧИВОСТЬ. Описание каждого гена начинается со сведений о размахе его изменчивости. Мы старались следовать принципу «матрёшек» - постепенно доставая из больших «матрёшек» содержащиеся в них всё более дробные регионы. В самых подробных описаниях мы начинали с размаха изменчивости в ойкумене, затем переходили к регионам Евразии, затем рассматривался размах изменчивости между субрегионами Северной Евразии, то есть бывшего СССР, далее - различия между народами Восточной Европы, а завершался маршрут самой маленькой матрёшкой: рассмотрением изменчивости между разными русскими популяциями. Обратим внимание на важный момент, который может вызвать недоумение. Порой различия между русскими популяциями достигают различий между регионами Евразии. Нет ли здесь ошибки? Нет. Просто надо давать себе всегда отчет, о каких частотах сейчас идет речь - усреднённых (региональных или этнических) или локальных (популяционных). Все региональные частоты и практически все этнические частоты представляют собой усреднённые величины. Например, частота какого-либо аллеля в Сибири представляет собой среднюю по всем изученным сибирским народам. Частота у коми - среднюю по всем изученным популяциям коми. При этом если частота в лишь какой-то одной популяции этноса была необычно высокой или низкой - на средней частоте аллеля в этносе это практически не скажется. И поэтому размах изменчивости между регионами - это различия между устойчивыми, усреднёнными региональными оценками. Для русских же популяций мы приводим размах изменчивости между локальными популяциями. И за счёт отклонений частоты всего в одной лишь популяции размах может стать большим. 3) КАРТА. Главная часть в любом описании - это рассмотрение карты. Сначала мы пытаемся сформулировать первое впечатление от карты - степень сложности рельефа и характер тренда. Затем переходим к деталям и отклонениям от основного увиденного тренда. Смотрим, каковы коэффициенты корреляции с географическими координатами, можно ли описать изменчивость гена в терминах «широтной» или «долготной» изменчивости. Чтобы оценить соответствие карты аллеля генетическому ландшафту всего генофонда, мы приводим и коэффициенты корреляции между данной картой и картами трёх главных компонент. В табл. 5.3.3. перечислены аллели, которые внесли наибольший вклад в главные сценарии генофонда - с коэффициентом корреляции выше 0.4. 4) ЧТО У СОСЕДЕЙ? А что происходит вокруг русского ареала? Эти данные мы берём уже не из карты, а из таблиц. Следует ли изменчивость в русском ареале более общим закономерностям, известным для данного аллеля? В частности, как варьирует частота аллеля у ближайших соседей - других восточноевропейских народов? Если на границе русского ареала частота начинает меняться, то какова частота у соседнего народа? Такое сравнение показывает, может ли это изменение генофонда в окраинных русских популяциях объясняться смешением с соседними народами. 5) ОТБОР. Каков уровень GST ДЛЯ данного аллеля, и как он соотносится со средним уровнем? Может быть, изменчивость данного аллеля сформирована под действием естественного отбора? Если отклонение от среднего уровня велико, то можно предположить действие стабилизирующего или же дифференцирующего отбора. 6) ГИПОТЕЗЫ. Какие ещё особенности видны на карте? Можно ли сформулировать гипотезу, объясняющую обнаруженный геногеографический рельеф? Как правило, нелегко дать объективную интерпретацию карты в терминах истории народа. Для этого необходим особый анализ, специальный инструментарий, совместная работа со специалистами в смежных областях - историками, археологами, этнографами, антропологами. Ассоциативное мышление здесь может сыграть злую шутку - чем оно богаче, чем больше мы знаем об этногенезе и этнической истории данного народа, тем проще любую особенность карты истолковать как древние или недавние миграции, следы прошедших племён и событий. Поэтому те немногие исторические экскурсы, которые мы включили в описания карт, следует рассматривать лишь как одну из множества гипотез. Этому плану - с теми или иными разумными отклонениями - мы следуем при описании каждого гена. И надеемся дать хотя и краткий, но достаточно объективный обзор его изменчивости. В конце концов, география отдельных генов - это непосредственная реальность, конкретное воплощение генофонда. Так рассмотрим же её со всем вниманием, ген за геном. И только уже после этого перейдём к самой структуре генофонда - той скрытой от непосредственного наблюдения реальности, которая формирует свои отдельные воплощения, а сама слеплена историей популяции. Её мы попробуем выявить в разделе 5.3., завершающем главу. 
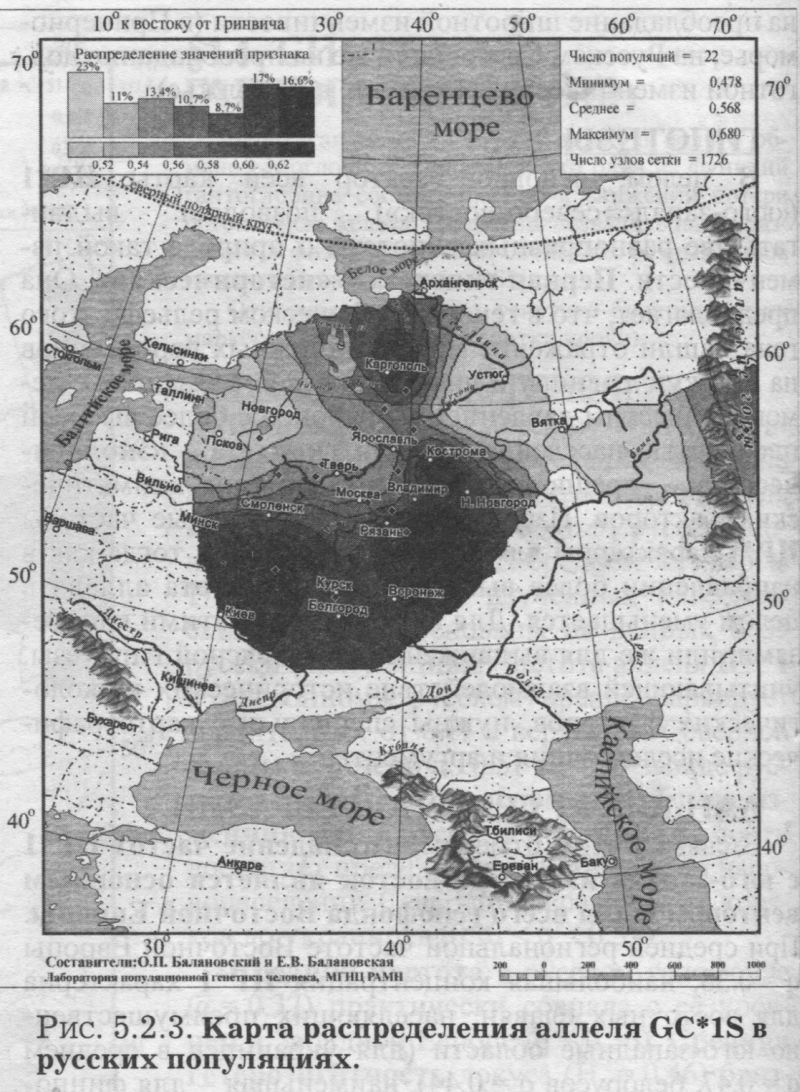
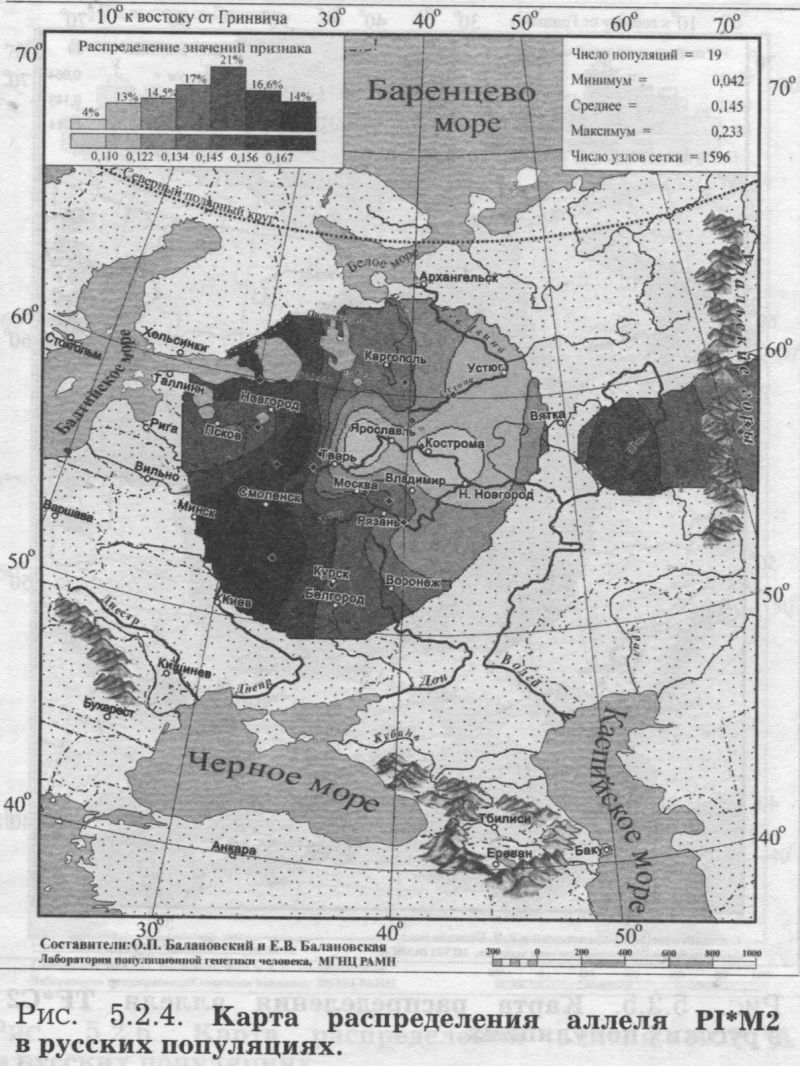
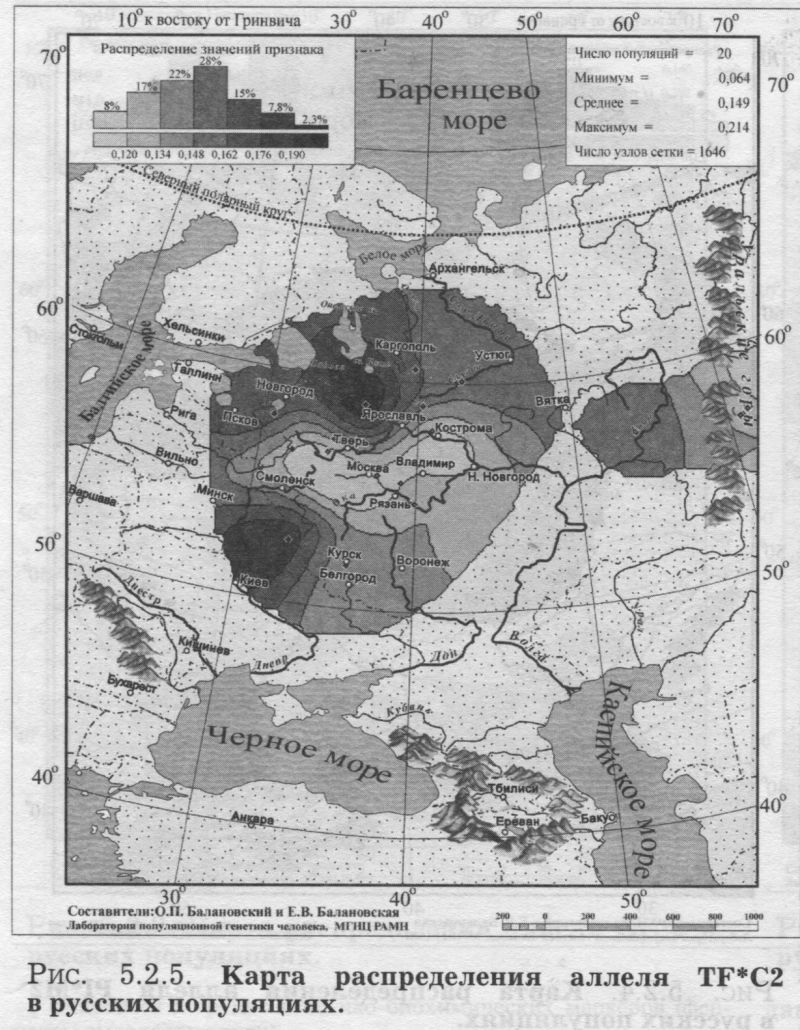
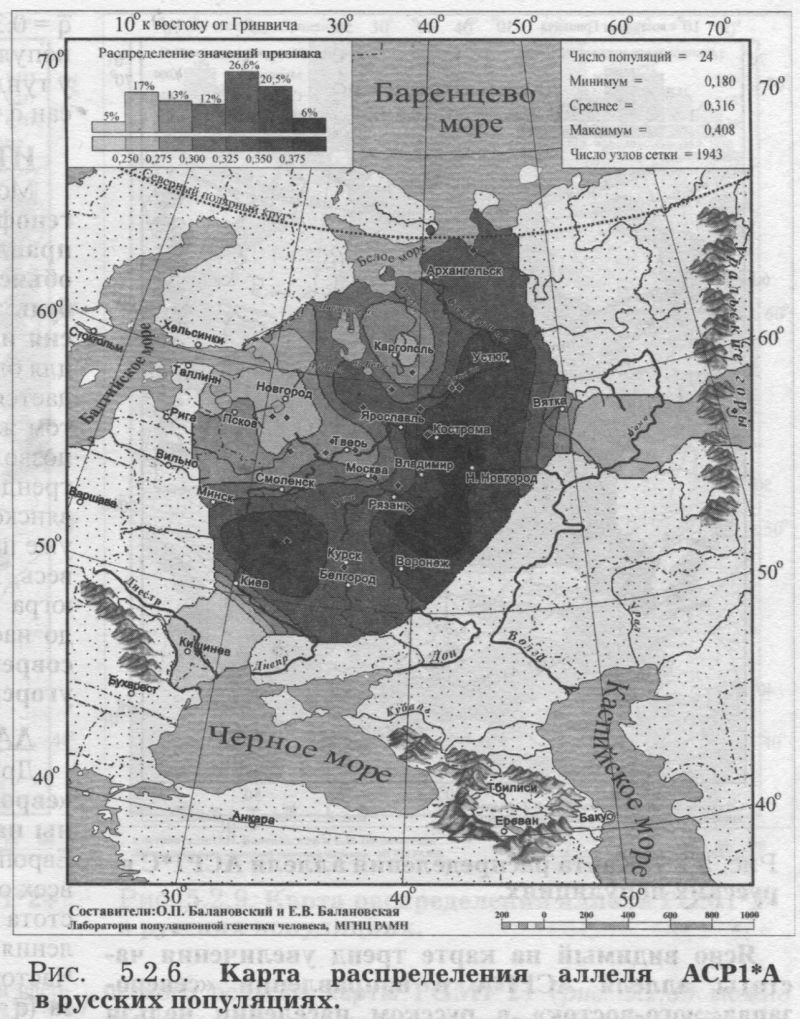
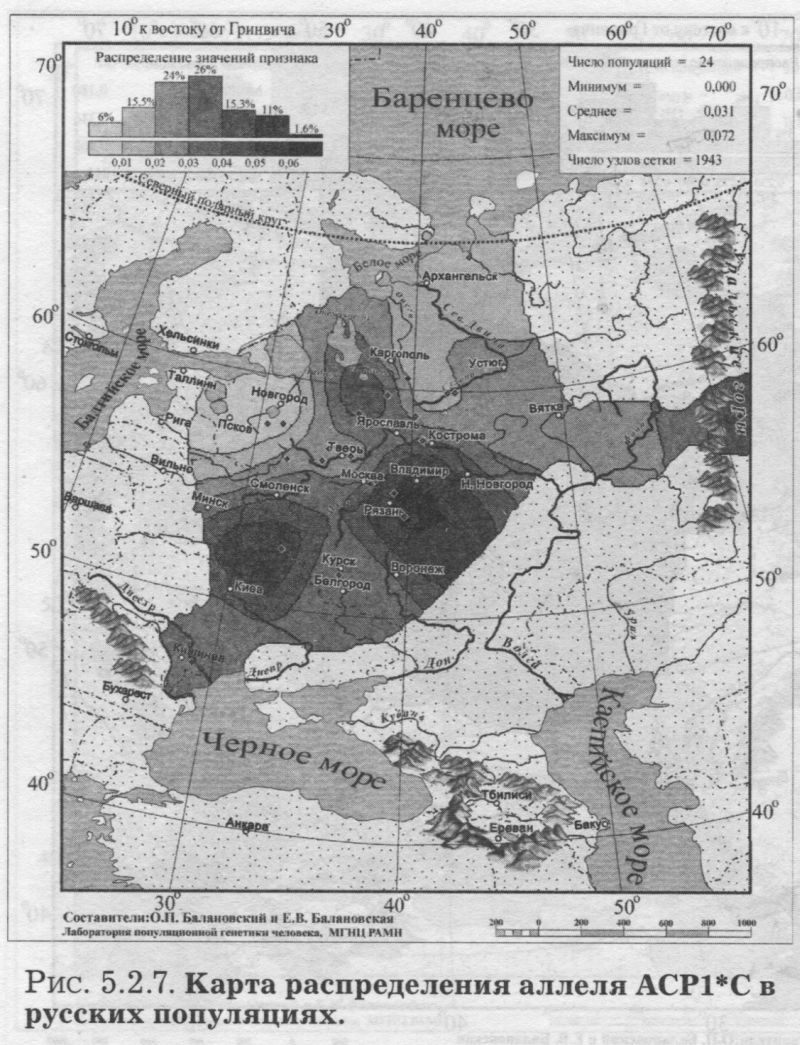
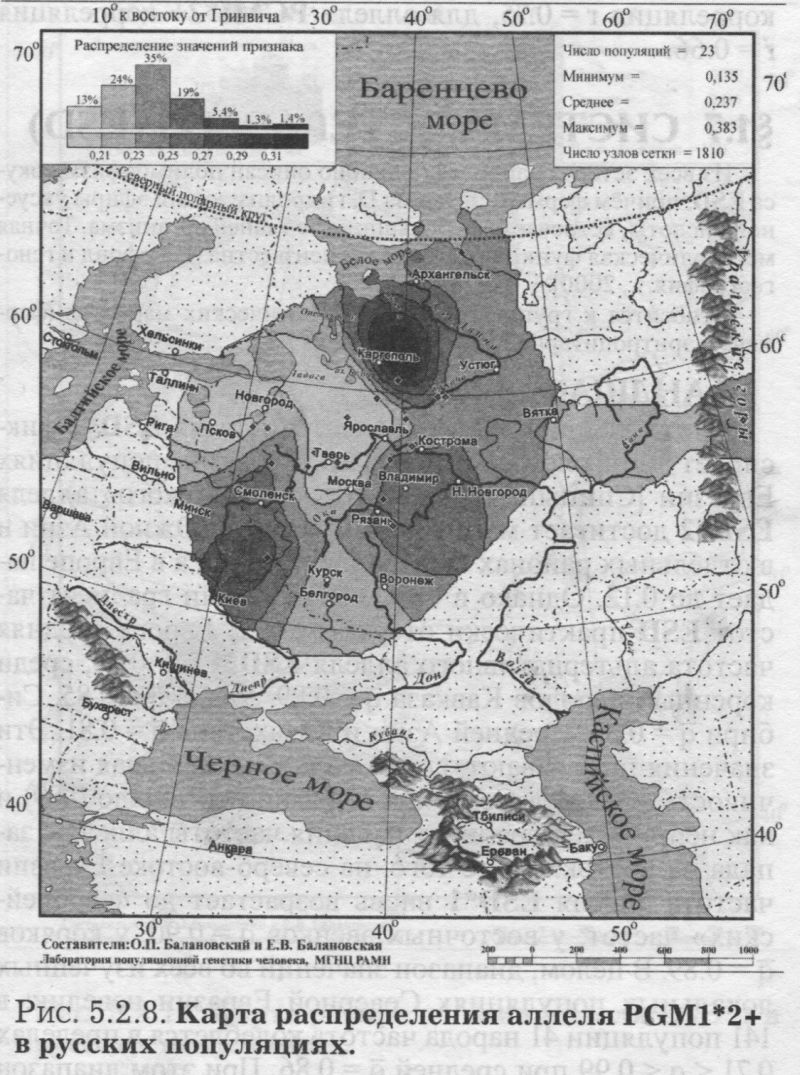
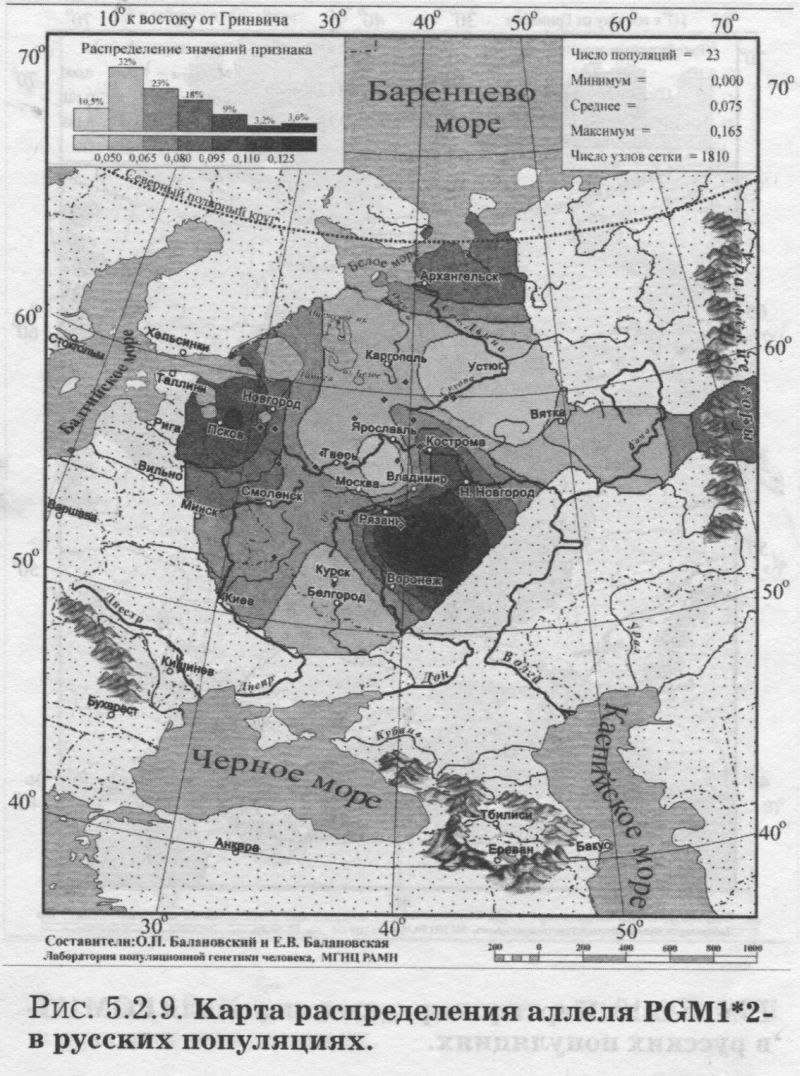
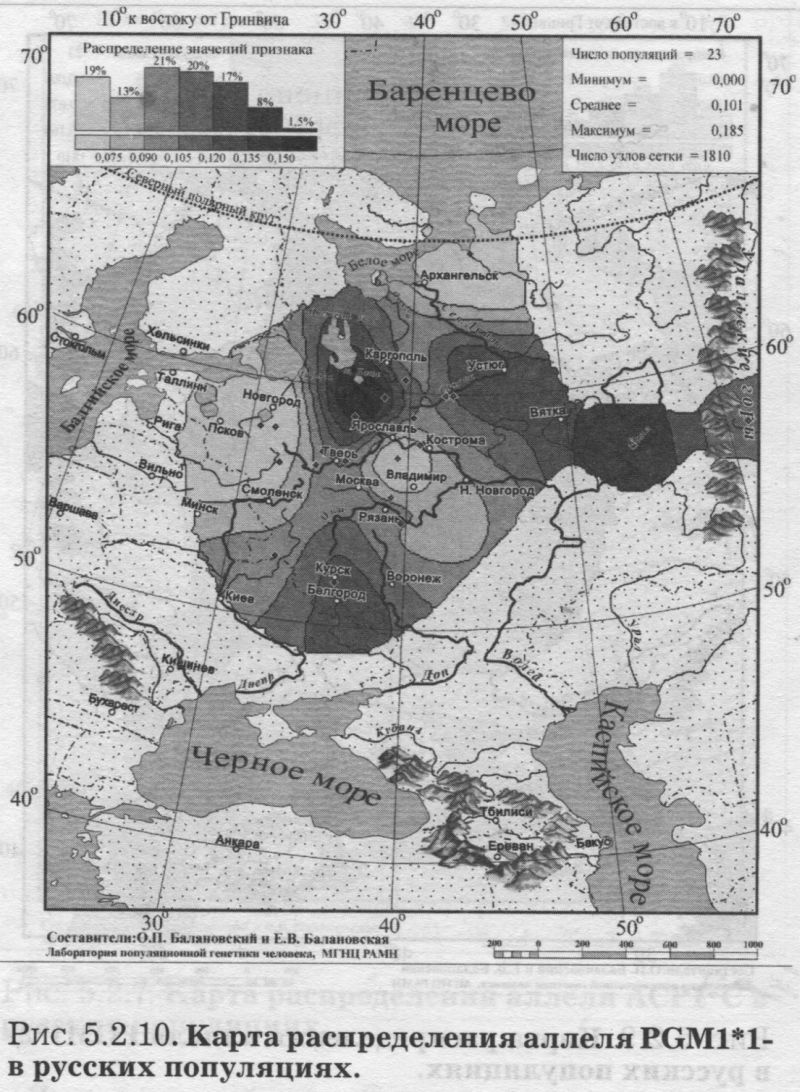
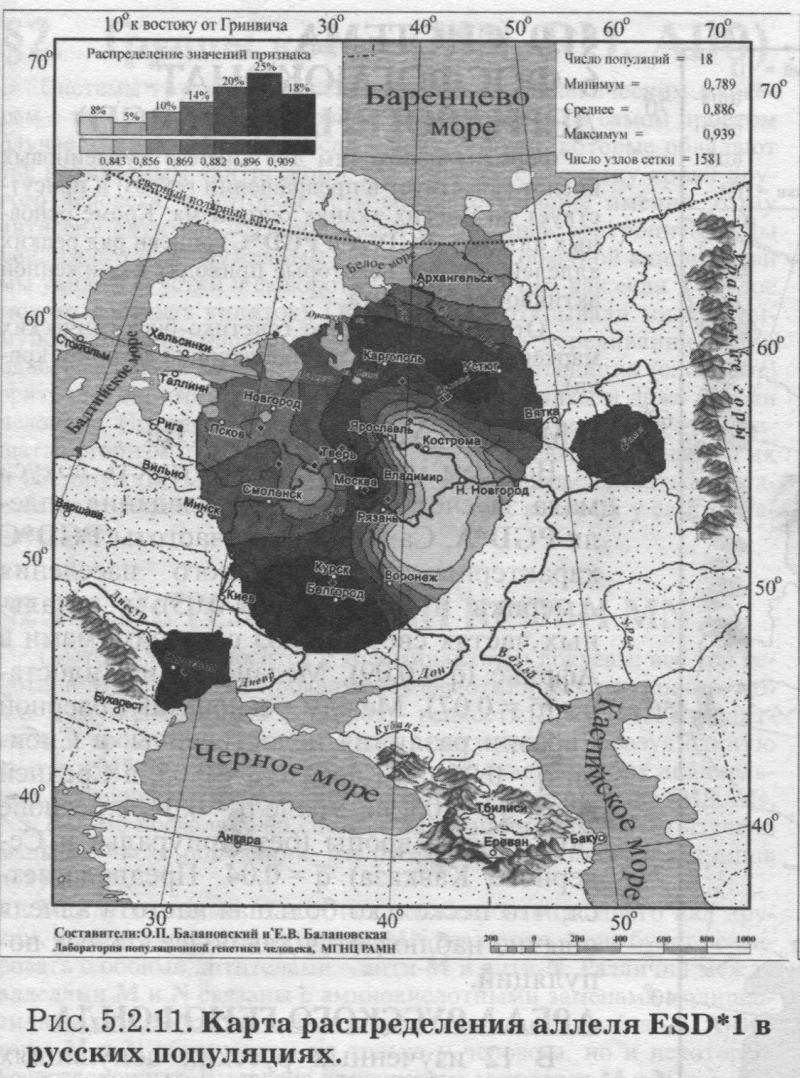
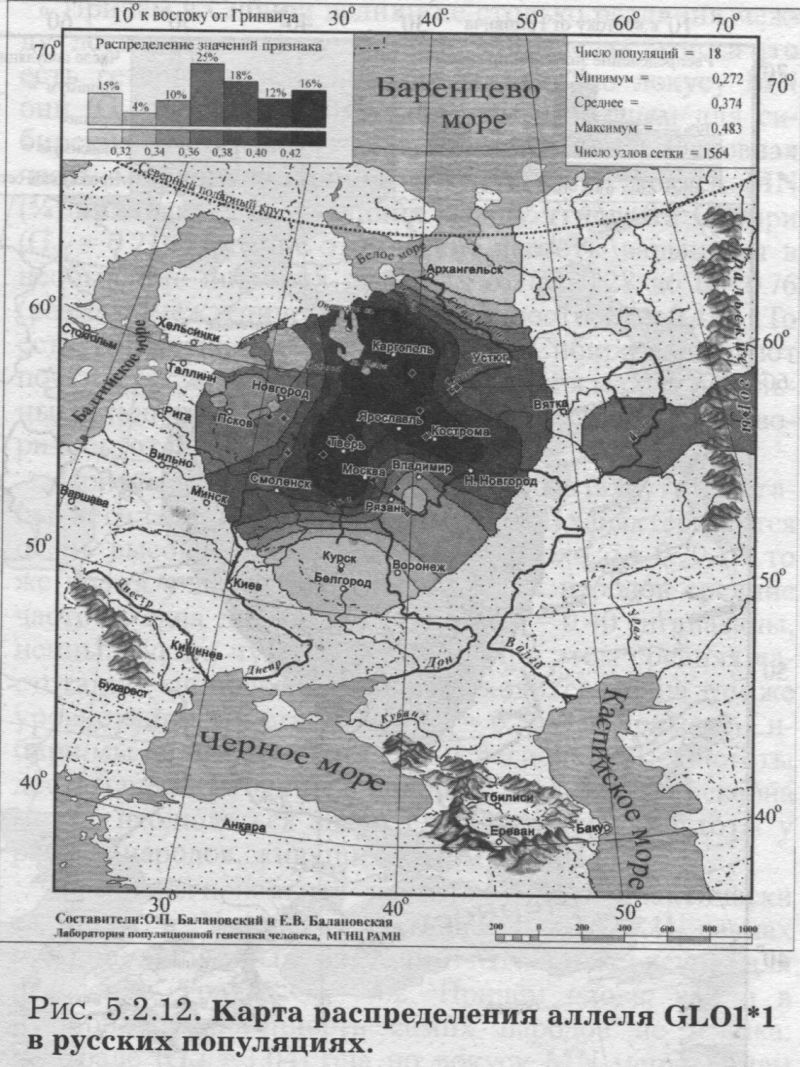
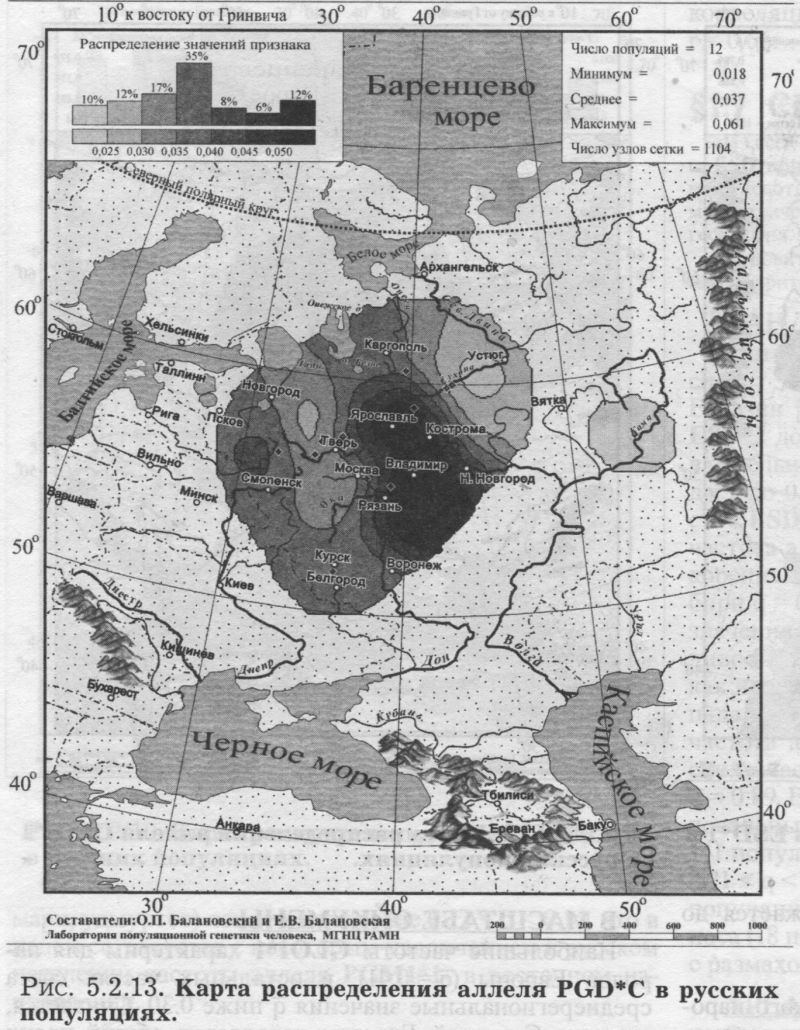
§1. Биохимические маркёры (HP, GC, PI, TF, ACPI, PGM1, ESD, GLOl, 6PGD)Сначала мы расскажем о ферментах сыворотки крови (HP, GC, PI, TF). а затем перейдем к эритроцитарным биохимическим маркёрам (AСР1, PGM1, ESD, GLOl, 6PGD) «Биохимическими» эти маркеры называются по методу их определения: ферменты (обычно крови) опознаются биохимическими методами, такими как электрофорез и изоэлектрофокусирование Упрощенно говоря, это выглядит так: на дорожке геля между катодом и анодом «разгоняется» специально подготовленный образец кровн Легкие молекулы бегут быстрее и обгоняют более тяжелые молекулы. В результате мы своими глазами видим, какой генотип у данного человека (ведь в биохимических локусах нет доминирования одного аллеля над другим). Например, если у него гстсрозигота по данному локусу, - мы увидим сразу две полоски на геле: одну для более «быстрого» аллеля (полученного им от одного из родителей), а вторую полоску для более «тяжелого» аллеля (полученного им от второго родителя). У гомозигот, получивших от обоих родителей одинаковые аллели, мы увидим, конечно, лишь одну полоску. Но. сравнив ее с «образцовой» матрицей, где примерно показано, как быстро должны «бегать» какие аллели, мы можем определить, какой именно аллель данного локуса имеется у данного человека. §1.1 СИСТЕМА ГАПТОГЛОБИНА (HP) Белок гаптоглобин играет важную роль в обмене гемоглобина. Полагают, что одна из функций гаптоглобина состоит именно в снижении потери организмом железа Отмечены незначительные различия в физиологических свойствах разных типов гаптоглобина [Спицын. 1985]. Относится к группе генетико-биохимических маркеров (ферменты сыворотки крови). АРГАЛ РУССКОГО ГГНОФОНЛА Полиморфизм системы гаптоглобина изучен в русском населении довольно подробно - карта распространения аллеля HР*1 основана на данных о 51 русской популяции (рис. 5.2.1.). В среднем же, карты 17 наиболее полно изученных классических маркёров основаны на данных о 35 русских популяциях. Частота аллеля HР*1 в «исконном» русском ареале варьирует значительно: от 0.17 в верховьях Северной Двины и Мезени до 0.51 на юго-западе русского ареала. Столь огромный размах изменчивости в русских популяциях почти достигает пределов вариации во всем народонаселении Северной Евразии - от коряков до украинцев (0.10 < q < 0.57). Средняя частота в русском генофонде (q = 0.37) практически совпала с её уровнем в Западной Европе (q = 0.36). Средняя гетерозиготность локуса (HS = 0.46) близка к максимально возможному значению для двухаллельного локуса (HS = 0.50). Межпопуляционное разнообразие карты (GST = 1.68) несколько выше среднего уровня дифференциации в русском генофонде (GST = 1.36)6. Таким образом, при общем рассмотрении вариации HP*1 в русском генофонде на первый план выходит типичность этого аллеля, его хорошая изученность и огромная изменчивость в русском ареале, что обязывает к внимательному рассмотрению геногеографии НР*1. Карта HP*1 (рис. 5.2.1.) выявляет изменчивость в русских популяциях, близкую к клиналыюй. Основной вектор изменчивости: иадение частоты аллеля в направлении «юго-запад<=>северо-восток» с заметным преобладанием широтной изменчивости над долготной. Этот вектор отражён и в коэффициентах корреляции с географическими координатами (теки. 5.2.1.): с широтой r = 0.427, с долготой r = 0.34. Причем в отдельных областях карты тот или другой коэффициент связи превышает уровень r = 0.8 (табл. 5.2.1.), указывая на преобладание широтной изменчивости (в Причерноморье, на Русском Севере) или же на преобладание долготной изменчивости (например, в Поволжье). ГИПОТЕЗЫ В целом, основной вектор всей карты НР*1 («юго-запад<=>северо-восток») позволяет выдвигать две равноправных гипотезы о природе такой изменчивости. Первая гипотеза - «историческая». Она предполагает, что в геногеографическом рельефе этого гена нашли отражение миграции южных европеоидов на Русскую равнину и диффузия генов, связанная с демографическим давлением из районов с более высокой плотностью населения. Вторая гипотеза - «экологическая» - предполагает прямое влияние климатических факторов. Известно, что максимальные частоты НР*1 характерны для тропического пояса, тогда как в направлении более высоких широт частота аллеля в целом уменьшается. Для выбора между этими гипотезами, или же для выдвижения комплексной гипотезы, учитывающей взаимодействие исторических и экологических факторов, нужны специальные картографические исследования и аргументы. ЛАНДШАФТ СЕВЕРНОЙ ЕВРАЗИИ В связи с этим отметим, что падение частот НР*1 с юго-запада на северо-восток является основным вектором и для всего генофонда Восточной Европы. При средней региональной частоте Восточной Европы q = 0.38, наибольшая концентрация НР*1 характерна для восточных славян, населяющих преимущественно юго-западные области (для украинцев в среднем q = 0.47, белорусов q = 0.44), наименьшая - для финноязычных народов северо-востока Европы - удмуртов, мари и коми-зырян (в среднем 0.31 < q < 0.34). Однако этот вектор не является общеевразийским. У манси (q = 0.40) и лесных ненцев (q = 0.41) Западной Сибири частота НР*1 вновь превышает отметку 0.4, а затем, после некоторого падения, вновь достигает в коренном населении Якутии «европейских» значений: в ряде популяций q>0.40 при средней q = 0.37. Карта распространения НР*1 в коренном населении Северной Евразии подробно проанализирована нами в работе [Балановская, Нурбаев, 1995]. В целом «хребет» высоких значений тянется от Украины до Якутии с пологим уменьшением частоты аллеля как в северном, так и в южном направлениях. Такой характер изменчивости нельзя свести ни к «европеоидно-монголоидным» взаимодействиям, ни к влиянию широтной изменчивости среды. РУССКИЙ СЕВЕР Чрезвычайно любопытно нарушение основного вектора на самом севере русского ареала (рис. 5.2.1). К северу от широкой зоны низких частот, включающей ядро минимальных значений НР*1 в бассейне Северной Двины и верховьях Мезени (q < 0.26), частота вновь возрастает до среднерусского уровня (q = 0.36). Эта локальная «арктическая» зона относительно повышенной частоты НР*1 расположена на побережье Баренцева моря, в низовьях Мезени и Печоры. В целом «арктическая» зона соответствует миграциям древнего новгородского населения, торговому пути в Мангазею и ареалу расселения поморов. Считается, что население этой территории сформировано преимущественно потомками новгородцев, а население более континентальных районов Русского Севера (в среднем и верхнем течении северных рек) сформировалось, главным образом, за счёт миграций из Ростово-Суздальской, а не Новгородской Руси. К сожалению, система гаптоглобина не позволяет дифференцировать новгородский и ростово-суздальский («низовой») миграционные потоки, поскольку сами новгородские и ростовские земли мало различаются по частотам НР*1. Сравнение с общей картиной северо-евразийского ландшафта НР*1 показывает, что повышение частоты в самых северных русских популяциях трудно связать с влиянием «субстратного» населения - поскольку в целом для коренного (дославянского) населения циркумполярной зоны характерны низкие частоты НР*1 (см. карты в работе [Балановская, Нурбаев, 1995]). Русские колонисты осваивали побережье интенсивнее, чем континентальные районы. Потому низкие частоты, выявленные в русских популяциях верховьев Северной Двины и Мезени, можно связать с большим участием «субстрата» (финно-угорских или самодийских племён) в формировании русского населения континентальных районов. Подобная трактовка изменчивости НР*1 на Русском Севере имеет ранг лишь предварительной гипотезы, и для корректных выводов необходимо более полное обследование ряда популяций. ИТОГ Итак, основная закономерность для НР*1 в русском населении - изменчивость, близкая к клинальной: с максимумом на юге и снижением частоты к северо-востоку. Общие закономерности ландшафта аллеля НР*1 в русском населении (включая отличия «арктической зоны») хорошо согласуются с первыми главными компонентами русского генофонда в целом: коэффициенты корреляции со всеми тремя первыми главными компонентами изменчивости классических маркёров варьируют в пределах 0.4 < г < 0.5. Хорошо они согласуются и с главными компонентами фенофонда русского народа по данным антропологии и дерматоглифики (глава 4). Это может означать, что система НР*1 высоко информативна при изучении структуры русского генофонда. §1.2 СИСТЕМА ВИТАМИН-D ТРАНСПОРТИРУЮЩЕГО Б£ЛКА (ГРУППОСПЕЦИФИЧЕСКИИ КОМПОНЕНТ GC) Предполагается, что основная функция группоспецифического компонента GC - связывание и транспорт витамина D3 и его метаболитов. Выделяются два основных аллеля GC*1 и GC*2, причём аллель с большей электрофоретической подвижностью подразделяется на два варианта - GC*1S и GC*1F [Генофонд и геногеография..., 2000]. Локус GC, локализованный на длинном плече 4-й хромосомы в районе сегментов qll-ql2, тесно сцеплен с локусом сывороточного альбумина. Есть указания, что некоторые различия в способности аллелей GC связывать витамин D приводят к их корреляции с факторами экологии и климата (тип питания, широта местности, высота над уровнем моря, интенсивность солнечной радиации, среднегодовая температура) [Спицын, 1985]. При этом частота аллеля GC*1 максимальна в наиболее тёмнопигментированных экваториальных группах населения, а в регионах с относительно низкой солнечной радиацией повышается частота GC*2. ЛАНДШАФТ ЕВРАЗИИ При сравнении населения основных регионов мира максимум частот GC*2 (q = 0.28) и GC*1S (q = 0.58) приходится на популяции Европы. Для Восточной Европы (без Приуралья и Северного Кавказа) частота GC*2 составила в среднем q = 0.31, слабо варьируя от q = 0.27 у литовцев до q = 0.33 у молдаван и украинцев. На Урале наблюдается общее понижение частоты аллеля GC*2, составляя в среднем q = 0.26. Средняя частота аллеля у уральских народов - башкир, чувашей, удмуртов, мари, коми, тундровых ненцев - также варьирует в узких пределах: от 0.25 до 0.28. Исключение составляют коми-пермяки, но они представлены лишь одной популяцией с частотой 0.38. В Зауралье у манси частота GC*2 понижается до q = 0.19. Среднерусские частоты аллелей GC*2 (q = 0.29 с вариацией от 0.14 до 0.38) и GC*1S (q = 0.57 с вариацией от 0.48 до 0.68) - совпадают с размахом изменчивости, характерным для населения всей Европы. Иными словами, частоты этих аллелей в русских популяциях отличаются друг от друга столь же существенно, как среднеэтннческие частоты у народов всей Европы - от Скандинавии до Испании! Для Северной Евразии в целом изменчивость GC намного выше, чем в Европе: частоты аллеля GC*2 варьируют от 0.05 до 0.77. Межпопуляционное разнообразие GC*2 (Gsr - 0.90) и GC*1S (GS| = 1.14) несколько ниже среднего для русского генофонда по всем классическим локусам уровня (Пет = 1 36), что может указывать на незначительное АРЕАЛ РУССКОГО ГЕНОФОНЛА Рассмотрим на этом евразийском фоне карты географической изменчивости аллелей СС*2 (рис. 5.2.2.) и GC*1S (рис. 5.2.3.), основанные на информации по 22 русским популяциям (К = 22). Генетический рельеф карт довольно сложен. Во многом они являются зеркальным отражением друг друга - ранговый коэффициент корреляции между картами составил r=-0.57. Однако для карты GC*2 (рис. 5.2.2.) наиболее характерным элементом является долготный «разлом» (пояс минимальных частот), следующий примерно по 40° долготы. Для карты GC1S (рис. 5.2.3.) наиболее характерна широтная изменчивость с повышением концентрации аллеля к югу от 55° параллели. При этом средняя корреляция частот обеих карт с долготой близка к нулю, а корреляция с широтой для аллеля GC*1S составила r = 0.30, достигая в отдельных районах r=0.85 (табл. 5.2.1). Самыми загадочными являются присутствующие на обеих картах диаметральные различия в частотах между соседними популяциями Вологодской области - бассейнов Онеги (GC*2, q = 0.17) и Сухоны (две популяции с частотами GC*2 q = 0.36 и q = 0.37). Отражают ли они реальные различия между генофондами этих популяции или же обусловлены выборочными эффектами (выборки составили 61, 64 и 163 индивида, соответственно), это покажут лишь дальнейшие генетические исследования Русского Севера ИТОГ Таким образом, нанойкуменный тренд (возрастание GC*2 к северу) не прослеживается в русском генофонде. Однако это предварительное заключение полностью зависит от достоверности низкой частоты этою аллеля на Онеге - если резкое понижение частоты GC*2 до 0.17 (и, соответственно, повышение частоты GC*1S) окажется случайностью и не подтвердится при дальнейших исследованиях, карты GC*2 и GC*1S в целом не будут противоречить гипотезе широтной изменчивости. Карга аллеля GC*1F высоко коррелирует с главным сценарием изменчивости классических маркёров в русском генофонде: r =0.63. Все три аллеля коррелируют со второй главной компонентой классических маркёров: для GC*1S г = 0.66; для GC*2 r = 0.43; для GC*1F r = 0.37. §1.3 СИСТЕМА ИНГИБИТОРА ПРОТЕАЗ (α1-АНТИТРИПСИНА PI)Белок обеспечивает до 90% всей активности ингибитора протсаз в отношении трипсина, ннгибнруя также сшс целый ряд ферментов - хнмотрнпенн, эластазу. коллагеназу. тромбин н протсазы лейкоцитов. О функциональной важности PI свидетельствуют достоверные ассоциации дефицита этого фермента с рядом заболеваний легких и печени. Система включает три субтнпа (РI*M1, Р1*М2, РI*М3 с суммарной частотой более 90%) основного варианта и значительное число редких вариантов Всс варианты PI функционально различны: уровень белка у гомозигот убывает в ряду M1>М2>М3, а редкие аллели продуцируют сшс меньшее количество белка [Генофонд и геногеография.... 2000]. Относится к группе генетнко-биохимических маркёров (ферменты сыворотки кровн). ЛАНДШАФТ ЕВРАЗИИ В населении Северной Евразии частота аллеля в целом убывает в направлении с запада на восток. Средняя частота Р1*М2 составила для коренных народов Кавказа q = 0.16, Урала - q = 0.14, Сибири q = 0.08. В народонаселении Европы частота аллеля РI*М2 убывает в ином направлении - с юга (q = 0.l8) на север (q = 0.09). Среднерусская частота аллеля РI*М2 (q = 0.15) находится в этом интервале. Среди 19 изученных русских популяций частота РI*М2 варьирует от 0.04 до 0.23. Однако межпопуляционное разнообразие РI*М2 (GST = 0.52) оказалось намного ниже среднего для русского генофонда уровня дифференциации (GST=1.36), что позволяет предполагать давление стабилизирующего отбора. АРЕАЛ РУССКОГО ГЕНОФОНДА Четкий градиент «запад<=>восток» (рис. 5.2.4.), наблюдаемый в Северной Евразии в целом, характерен и для русского генофонда (корреляция частоты РI*М2 с долготой достигает в отдельных районах значений г = 0.9). Однако среднее значение коэффициента корреляции с долготой невысоко (г=0.30), поскольку на самом востоке карты, у русских популяций Урала, вновь наблюдается резкое повышение частоты аллеля Карты всех трёх аллелей обнаруживают очень высокую корреляцию со вторым сценарием изменчивости русского генофонда по классическим маркёрам: для РI*М1 коэффициент связи достигает чрезвычайно высокого значения r = 0.87; для РI*М2 r = 0.68; для РI*МЗ r = 0.64. ПОПУЛЯЦИОННЫЙ ЭКСПЕРИМЕНТ: ВКЛЮЧАEМ ГОРОДСКОЕ НАСЕЛЕНИЕ. Не только на рассмотренных, но и на многих других картах (PI*S, GC*1S, PGM1*1-, PGM1*2-, TF*C2, TF*DCHI, ACP*A) в русских популяциях Урала также ярко видны резкие нарушения тех трендов, которые характерны для «исконного» русского ареала. Напомним, что в порядке эксперимента в анализ были включены городские популяции, лежащие за пределами «исконного» русского ареала. Например, для локуса PI это русское население республики Удмуртия, изученное в г. Воткинске (где частота РI*М2 q = 0.16, в то время как для самих удмуртов характерна средняя частота q = 0.10), Екатеринбурга (с частотой PI*M2 q = 0.15) и Асбеста (с частотой PI*M2 q = 0.14). Эти значения лишь воспроизводят (с незначительной вариацией) среднерусскую частоту (q = 0.15) и не несут никакой дополнительной информации ни о географии, ни об истории русского генофонда. Городское русское население Урала сформировано, главным образом, в результате исторически недавних миграций. И потому эти значения можно не изучать: их можно просто прогнозировать, исходя из демографических данных о миграционных потоках нз «исконного» русского ареала. Таким образом, проведённый эксперимент подтверждает правильность нашей исходной позиции: население, находящееся за пределами «исконного» ареала народа (и тем более городское!), не информативно для изучения пространственной структуры генофонда. Все усилия целесообразно направить на выявление генетической изменчивости сельских популяций в пределах «исконного» ареала. Зная генофонд «исконного» ареала, зная источники, направление и интенсивность миграционных потоков (откуда и сколько мигрантов прибыло в данную популяцию за пределами «исконного» ареала), мы можем с большой степенью вероятности прогнозировать генетический состав исторически «молодых» популяций. §1.4 СИСТЕМА ТРАНСФЕРРИНА (TF)Трансферрин образует соединение с железом, нейтрализуя токсичные ионы, и транспортирует его в костный мозг. Множество вариантов трансферрина контролируется тремя сериями аллелей - TF*B, TF*C, TF*d, каждая из которых включает несколько аллелей. Относится к группе генетико-биохимических маркёров (ферменты сыворотки крови). РУССКИЕ ПОПУЛЯЦИИ Из редких вариантов всегда рассматривается аллель TF*DCHI, поскольку увеличение его частоты традиционно интерпретируется как признак монголоидной примеси [Спицын, 1985; Шнейдер, 1999]. Однако средняя частота TF*DCHI (табл. 5.2.1.) в русском населении находится ниже даже 1%критерия полиморфизма (q = 0.009). Максимальные значения 7(f в основном ареале обнаружены в популяциях Рязанской, Костромской и Вологодской областей (qMAX = 0016) при среднем размере выборки менее 100 человек, которая слишком мала для надёжного тестирования столь редкого аллеля. Поэтому, не приводя карту TF*DCHI, лишь укажем, что она намечает слабую тенденцию увеличения частоты TF*DCHI с запада на восток, однако достоверность такого тренда невелика. Из пяти карг TF, построенных для русскою генофонда (их характеристики даны в таСа. 5.2.1.), приведем карту аллеля TF*C2 (рис. 5.2.5.), созданную по данным о 20 русских популяциях. Частота TF*C2 варьирует от 0.06 до 0.21 со среднерусским значением q = 0.15. Гетерозиготность локуса (по совокупности всех аллелей) HS = 0.365. Важно отметить, что межпопуляционное разнообразие других аллелей локуса значительно превышает среднерусский уровень изменчивости (GST=1.36): для TF*B01 , для TF*C3 GST=2.05. Это позволяет выдвигать гипотезу подверженности аллелей TF*B01 и TF*C3 дифференцирующему отбору. Однако для аллеля TF*C2 характерно резко сниженное межпопуляционное разнообразие: GST = 0.47. Достоверно сниженный размах изменчивости TF*C2 может указывать на давление стабилизирующего отбора на этот вариант трансферрина. ЛАНДШАФТ ЕВРАЗИИ Хотя частота TF*C2 возрастает до 0.40 в некоторых популяциях Южной Азии, но и для популяций Европы, и для коренных народов Приуралья она лежит примерно в одном диапазоне частот (от 0.07 до 0.27), причём среднеэтнические частоты одинаковы у самых разных народов: 0.10 - и у чувашей, и у финнов; 0.14 - и у мордвы, и у немцев; 0.15 - и у русских, и у испанцев; 0.16-0.18 - у мари, итальянцев, шведов и греков. АРЕАЛ РУССКОГО ГЕНОФОНДА Но, несмотря на такое мозаичное распределение на европейском пространстве, в русском ареале карта являет вполне выраженнуюзакономерпость: увеличение частоты TF*C2 на севере (0.16 < q < 0.21) и уменьшения (0.06 • q • 0.12) - в центральной части (волго-окском междуречье). Граница проходит примерно по широте Твери. Что же касается «юго-западного» ядра высоких значений, то оно связано лишь с одной популяцией - староверов Сибири, искусственно помещённых на карте в Брянскую область по месту происхождения этой популяции [Спицын и др., 2001]. Очевидно, такая популяция не может служить надёжным показателем. Остальные изученные южные популяции - Рязани и Курска - характеризуются среднерусскими частотами (0.14 и 0.15, соответственно). ИТОГ Таким образом, на карте распространения аллеля TF*C2 в русском ареале можно выделить три широтных зоны: северная - с высокими частотами TF*C2, центральная - с пониженными частотами TF*C2, южная - со среднерусскими частотами. Карты всех трёх вариантов аллеля TF*C обнаруживают высокую корреляцию с первым сценарием изменчивости классических маркёров: для TF*C1 коэффициент связи достигает чрезвычайно высокого значения r = 0.80; для TF*C2 r - 0.58; для TF*C3 r - 0.57. Карта аллеля TF*DCHI, в свою очередь, высоко коррелирует с третьей главной компонентой русского генофонда: r = 0.79. А аллель TF*B вносит определённый вклад (r ≈ 0.4) и в первый, и во второй главные сценарии русского генофонда. §1.5 СИСТЕМА ЭРИТРОЦИТАРНОИ КИСЛОЙ ФОСФАТАЗЫ (ACPI)Предполагается, что основная функция ACPI состоит в регуляции внутриклеточной концентрации флавиновых ферментов и коферментов Кроме трех основных аллелей АСР1*А, АСР1*В, АСР1*С. существует ряд редких вариантов ACPI Все фенотипы различаются по средней активности фермента, причем количественные эффекты всех аллелей образуют ряд: А < В < С Относится к группе генетико-биохнмических маркёров (ферменты эрнтроцитарнон фракции крови). В МАСШТАБЕ ОЙКУМЕНЫ (АСР1*А) Для аллеля АСР1*А в генофонде ойкумены отмечается положительная связь частоты с географической широтой |Спицын, 1985; Spitsyn et al., 1998]. Однако для коренного населения Северной Евразии характерна совершенно иная и своеобразная долготная изменчивость эюго аллеля. Гряда относительно высоких значений АСР1*А (q>0.40), протянувшаяся от Алтая на север, отделяет западное «плато» средних значений АСР1*А от «низин» Средней Сибири и Саян (области низких частот АСР1*А в интервале 0.17 < q < 0.30). Далее на северо-восток частота аллеля вновь растёт до максимальных евразийских величин (0.55 < q < 0.62 у коряков, чукчей и эскимосов). На другом конце континента, на западе евразийского ареала, средняя частота АСР1*А составила q = 0.32 (в населении Западной Европы). Г>га величина совпадает и со среднерусской частотой q = 0.32, и со средней частотой аллеля у коренных народов Урала (q = 0.32) и Кавказа (q = 0.31). Лишь для народов Средней Азии наблюдается понижение средней частоты АСР1*А (q = 0.28) [Балановская, 1998]. АРЕАЛ РУССКОГО ГЕНОФОНДА (АСР1*А) Карта изменчивости частоты этого аллеля (АСР1*А) в русском населении, построенная по информации о 24 русских популяциях (К = 24), приведена на рис. 5.2.6. Она показывает, что для русского населения (так же, как и для населения Северной Евразии в целом) не характерно возрастание частоты аллеля в северных широтах, связываемое с широтной климатической зональностью, с возможным действием отбора. К тому же и уровень межпопуляционной изменчивости (GST = 1.28) соответствует селективно-нейтральному (HS = 1.36), и тем самым противоречит гипотезе отбора по этому аллелю. Не сказывается и влияние азиатских степей. Поскольку среднеазиатские популяции характеризуются снижением частоты аллеля, то их влияние должно было бы сказаться в убывании частоты АСР1*А в юго-восточном направлении. Например, ближайшие тюркоязычные популяции характеризуются следующими частотами АСР1*А: у казахов q = 0.26, у башкир q = 0.31, у татар q = 0.33, у чувашей q = 0.25 [Генофонд и геногеография ..., 2000]. Однако карта демонстрирует обратный тренд - не убывание, а возрастание в русском ареале частоты аллеля с северо-запада на юг и юго-восток. При этом вариации частот АСР1*А достаточно велики - от 0.18 до 0.41 (табл. 5.2.1.). Северо-западные русские популяции характеризуются сниженными частотами с минимумами в землях новгородских (q = 0.25), псковских, тверских, затем в Поонежье и далее к Архангельску. Высокие частоты характерны для востока и юго-востока с максимумами в землях костромских и рязанских (q >0.40). Популяции, выходящие за пределы основного ареала (Украина, Урал), демонстрируют частоты аллеля, характеризующие не столько структуру генофонда, сколько структуру поздних переселений (этот вопрос обсуждался в подразделе § I.3.). Эти «внешние» (по отношению к «исконному» русскому ареалу) популяции меняют тренд на противоположный и приводят к резкому снижению средних коэффициентов корреляции как с широтой, так и с долготой местности (табл. 5.2.1). Ясно видимый на карте тренд увеличения частоты аллеля АСР1*А в направлении «северо- запад<=>юго-восток» в русском населении нелыя объяснить, исходя из данных о его соседях. Например, у литовцев средняя частота АСР1*А составила q = 0.34, а у молдаван q = 0.25 [Генофонд и геногеография..., 2000], то есть у западных соседей наблюдается падение частоты аллеля с севера на юг - в противоположность тренду, наблюдаемому в русских популяциях - увеличения частоты аллеля с севера на юг. Поскольку данные по украинцам и белорусам отсутствуют, приведем средние частоты АСР1*А среди других славянских народов: у поляков q = 0.35, у чехов q = 0.32, у хорватов q = 0.28, у болгар q = 0.16. Мы вновь видим общеевропейский градиент падения частоты аллеля с севера на юг, противоположный наблюдаемому тренду в русском генофонде. В любом случае, данные о славянских народах не дают удовлетворительного объяснения тренду, выявляемому картой (рис. 5.2.6.). ГИПОТЕЗЫ (АСР1*А) Можно, однако, выдвинуть гипотезу о том, что в выявленном «русском» тренде АСР1*А нашли отражение генетические характеристики финно-угорского субстрата - тех древних насельников Русской равнины, которые освоили её задолго до того, как она стала Русской: до прихода славянских племён. На возможность такой гипотезы указывает тенденция некоторого увеличения частоты аллеля с севера и запада к югу в современных финно-язычных популяциях: у карел q = 0.24, коми-зырян q = 0.30, у коми-пермяков q = 0.30, у удмуртов q = 0.30, у мари q = 0.36, у мордвы q = 0.39. На севере, в самодийских популяциях, частота аллеля в среднем также невелика: у тундровых ненцев q = 0.31, у лесных q = 0.16, у нганасан q = 0.29. ИТОГ (ACP1*A) Можно резюмировать, что наблюдаемый в русском генофонде тренд роста частоты аллеля АСР1*А в направлении «северо-западоюго-восток» не поддастся объяснению в плане «глобальных» взаимодействий, будь то панойкуменный тренд, славянская экспансия или влияние евразийских степей. Вместе с тем, для ближайших финно-угорских народов также наблюдается нарастание частоты аллеля приблизительно в том же направлении: с севера (и с запада) на юг. Это позволяет предположить, что формирование данного тренда относится ко времени, предшествовавшему славянской экспансии на Восточно-Европейскую равнину, уже населенную финно-угорскими племенами (корела. весь, меря, мурома, чудь Заволочская, пермь, печера, югра и многими другими, имена которых не дошли до нас). Выявленный тренд, возможно, отражает в современном русском населении древний финно-угорский субстрат. ЛАНДШАФТ ЕВРАЗИИ (АСР1*С) Другой аллель - АСР1*С - принято называть «европейским». Для населения Европы характерны наибольшие частоты АСР1*С: средняя частота в Европе q = 0.05, в то время как в коренном населении всех остальных частей света - меньше 1%. Средняя частота АСР1*С составила q = 0.005 для коренного населения как Сибири, так и Средней Азии и Казахстана. Частота АСР1*С несколько выше в населении Кавказа (q = 0.016) и ещё выше у коренных народов Урала (q = 0.028). составляя у мордвы, мари, коми-пермяков и коми-зырян 4-5%. АРЕАЛ РУССКОГО ГЕНОФОНДА (АСР1*С) Карта аллеля АСР1*С (рис. 5.2.7.) демонстрирует в русском населении несколько иной характер изменчивости, чем предыдущая карта АСР1*А. Можно видеть общее снижение частоты аллеля к северу от волго-окского междуречья (до 1-2%) и повышение частоты АСР1*С в центральных, восточных и южных областях (до 7%). Однако, хотя корреляция с широтой и составила в среднем r = 0.34, для достоверной оценки различий при низких частотах (со средней q = 0.03) требуются большие выборки. Карты всех трёх аллелей локуса ACPI обнаруживают высокую корреляцию (0.6 §1.6 СИСТЕМА ФОСФОГЛЮКОМУТАЗЫ 1 (PGM1)Фермент фосфоглюкомутаза играет важную роль в метаболизме углеводов Из серии аллелей локуса PGM1 наиболее полно изучено распространение частых аллелей PGM1*1+, PGM1*1-, РGМ1*2+, РGМ1*2-. Относится к группе генетико-биохимических маркеров (ферменты эритроцитарной фракции крови). ЛАНДШАФТ ЕВРАЗИИ (РGМ1*1+, РGМ1*2+) Концентрация наиболее частого аллеля PGM1*1+ (его средняя частота в русских популяциях q = 0.59) растёт в Северной Евразии в направлении с запада на восток. Однако для русского населения этот градиент не прослеживается. Впрочем, то же можно сказать и об остальных аллелях локуса (рис. 5.2.8.-5.2.10.). Например, для аллеля PGM1*2+ в населении Евразии отмечается выраженный долготный градиент увеличения частоты в западном направлении: средняя частота q = 0.18 характерна и для коренного населения Сибири, и для Средней Азии; выше средняя частота в населении Урала (q = 0.23), и ещё выше у коренных народов Кавказа (q = 0.27). Однако в изменчивости русского народа этот долготный градиент PGM1*2+ проявляется нечетко. АРЕАЛ РУССКОГО ГЕНОФОНДА Этот, второй по частоте аллель PGM1*2+ варьирует в русских популяциях от 0.14 до 0.38 со средним значением q = 0.24 (табл. 5.2.1.). Но максимум частоты приходится па одну нз шести изученных популяций Вологодской области. Такой скачок частоты вероятнее всего связан с особенностями выборки. Ведь в среднем по шести вологодским популяциям частота PGM1*2+ (q = 0.23) неотличима от среднерусской (q = 0.24). К тому же и объем выборки, давшей максимальную частоту, невелик: n = 61 человек. Так что единственным важным структурообразующим элементом генетического рельефа карты PGM1*2+ (рис. 5.2.8.) можно считать снижение частоты аллеля на северо-западе, в землях новгородских и тверских. Частоты третьего аллеля PGM1*2 - варьируют в пределах 0 < q < 0.16 со средним значением q = 0.075. Карта аллеля PGM1*2- (рис. 5.2.9.) выявляет независимое повышение его частоты в трёх удалённых друг от друга популяциях - новгородской, архангельской и рязанской. Эти ядра высоких значений хорошо видны на фоне широкой области низких частот в северной половине русского ареала, вновь демонстрируя мозаичность генетического рельефа. Карта этого аллеля (PGM1*2-, (рис. 5.2.9.) во многих деталях является зеркальным отражением изменчивости карты аллеля PGM1*1- (рис. 5.2.1(1). Коэффициент ранговой корреляции между этими двумя картами составил r = - 0.62. Частоты четвертого аллеля PGM1*1- (рис. 5.2.10.) варьируют в тех же пределах: от 0 до 0.18 со средним значением q = 0.10 (табл. 5.2.1.). Однако некоторую закономерность в генетическом рельефе аллеля PGM1*1- всё же можно предположить. В целом средние и низкие частоты характерны для западных (новгородско-псковских) и волго-окских популяций. Высокие частоты наблюдаются к северу и востоку - в вологодских и костромской популяциях. Нарушают такую широтную изменчивость увеличения частоты аллеля PGM1*1- с юга на север лишь две географически крайних популяции - Курская (q = 0.14: n = 200 чел.) и Архангельская (q = 0.05: n = 96 чел.). Средние частоты соседних финноязычных народов Приуралья (мордвы, мари, удмуртов) лежат в интервале 0.12 < q < 0.14, что в целом согласуется с некоторым увеличением в русском населения частоты аллеля PGM1*1- в восточном направлении. ИТОГ Средний по всем аллелям локуса уровень межпопуляционной изменчивости (GST = 0.75) почти в два раза меньше селективно-нейтральной дифференциации (GST = 1.36), что может указывать на действие стабилизирующего отбора. Однако картина изменчивости аллелей столь мозаична, что её сложно интерпретировать как в терминах истории, так и в терминах отбора. Лишь дальнейшее изучение может выявить реально существующую генетическую изменчивость русского генофонда - 23 популяции, изученные по локусу PGM1, не позволяют выдвинуть сколько-нибудь информативную гипотезу о факторах, сформировавших генетический рельеф его аллелей. В целом можно предполагать, что информативность локуса PGM1 для изучения структуры русского генофонда невелика. Тем не менее, даже в изменчивости локуса PGM1 проглядывают общие закономерности архитектоники русского генофонда. Эти черты мы обнаруживаем, анализируя корреляции между картами аллелей локуса PG.M1 и каргами главных компонет русского генофонда (maat. 5.3.3.). Карта аллеля PGM1*2- обнаруживает высокую корреляцию (r = 0.76) с первым сценарием изменчивости классических маркёров. Два других аллеля вносят весомый вклад в изменчивость третьей главной компоненты русского генофонда: для аллеля PGM1*1+ корреляция r=0.75, для аллеля PGM1*2+ корреляция r = 0.66. §1.7 СИСТЕМА ЭСТЕРАЗЫ D (ESD)Из всех эстераз наиболее подробно описан полиморфизм локуса ESD. причём фермент эстераза D, гидролизующий эфиры уксусной кислоты, встречается в большинстве тканей организма. Точная метаболическая функция эстеразы D неизвестна [Генофонд и геногеография..., 2000). Относится к группе генетико-биохимических маркёров (ферменты эритроцитарной фракции крови). ЛАНДШАФТ ЕВРАЗИИ Считается, что градиент частот аллеля ESD*2 фиксирует монголоидный генный поток в популяциях Евразии [Спицын, 1985]: частота «азиатского» аллеля ESD*2 достигает максимума (q>0.40) в Южной Азии и в отдельных районах 11ентральной Азии, а в Европе падает до 0.12. Однако в Северной Евразии градиент частот ESD практически отсутствует. В Европе средняя частота альтернативного аллеля ESD*1 q = 0.88, среди коренных народов Кавказа q = 0.89, Урала q = 0.85, Сибири q = 0.83, Средней Азии и Казахстана q = 0.81. Эти значения показывают, сколь мала региональная изменчивость ESD*1 в Северной Евразии (от 0.81 до 0.89) и как неприметен градиент падения частоты аллеля с запада на восток. Более того, на северо-востоке Евразии частота аллеля ESD*1 вновь возрастает до «европейских» частот: у восточных эвенков q = 0.90, у коряков q = 0.89. В целом, диапазон значений во всех изученных локальных популяциях Северной Евразии невелик: в 141 популяции 41 народа частота колеблется в пределах 0.71 < q < 0.99 при средней q = 0.86. При этом диапазон популяционной изменчивости в пределах русского этноса (18 популяций, 0.79 < q < 0.94, q = 0.89) сопоставим с размахом изменчивости во всей Северной Евразии. АРЕАЛ РУССКОГО ГЕНОФОНДА Столь слабо выраженные межрегиональные различия не позволяют уверенно интерпретировать генетический рельеф ESD*1 (рис. 5.2.11.) в русском ареале как следы тех или иных миграций. А такое предположение возникает при первом же взгляде на карту - её рельеф как бы задаётся резко выраженной впадиной низких значений на юго-востоке, ограниченной с севера и юга грядами наиболее высоких частот ESD*1. Естественно предположить, что уменьшение частоты в юго-восточной части русского ареала связано с влиянием степных номадов Однако это предположение маловероятно, поскольку у ближайших тюркоязычных народов, изученных по этому гену, наблюдаются довольно высокие средние частоты ESD1*1: у башкир (q = 0.88) и татар (q = 0.88). У других восточных соседей средние частоты аллеля варьируют следующим образом (двигаясь с юга на север): у мордвы q = 0.89, у чувашей q = 0.81, у удмуртов q = 0.88, у коми-пермяков q = 0.83, у коми-зырян q = 0.88. На западе у карел частота ESD*1 достигает q = 0.91. На карте ядро относительно низких частот образовано только двумя русскими популяциями Костромы (q = 0.79, n = 76 чел.) и Рязани (q = 0.81, n = 76 чел ). При этом в Курской области и Волго-Окском междуречье частота ESD*1 возрастает до 0.90, а в новгородских и архангельских землях снижается до 0.86. ИТОГ Вновь карта генетического рельефа русского народа отражает не «европеоидно-монголондные» взаимодействия, в терминах которых привычно осмысливать генетическую изменчивость русских популяций, а сложные пути формирования русского генофонда. Объективно мы видим на карте чередование зон - широтных «волн», незначительно отличающихся то повышением, то понижением частоты аллеля. При этом средняя частота аллеля ESD*1 (q = 0.89, число популяций К = 18) полностью соответствует европейским стандартам, а межпопуляционная изменчивость (GST = 0.95) несколько ниже селективно-нейтрального уровня (GST = 1.36). Корреляции карг локуса ESD с картами главных компонент русского генофонда (табл. 5.3.3.) обнаруживают высокую корреляцию с первым (r = 0.58) и с третьим (r = 0.51) сценариями изменчивости классических маркеров. §1.8 СИСТЕМА ГЛИОКСАЛАЗЫ 1 (GLO1)Локус контролирует одну нз форм глноксалазы. отвечающей за превращения мстилглноксаля в лактат. Известны три варианта GLOl с активностью фермента, убывающей в ряду: GL01*2>GL01*1>GL01*0 («молчащий» аллель) Относится к группе генетико-биохимических маркёров (ферменты эритроцитарной фракции крови). В МАСШТАБЕ ОЙКУМЕНЫ Наибольшие частоты GLO1*1 характерны для пародов Европы (q = 0.41), в остальных частях света среднерегиональные значения q ниже 0.30. Считается, что для Северной Евразии характерен общий тренд увеличения частоты GLO1*1 с северо-востока на запад: диапазон изменчивости GL01*1 в 147 популяциях 44 этносов Северной Евразии составил 0.08 < q < 0.58 (q = 0.36). Однако максимальные частоты наблюдаются не только в центральных и северных районах европейской части России, по и па севере Западной Сибири, где частота аллеля возрастает до 0.51, приближаясь к мировому максимуму. Однако в среднем народы Сибири (q = 0.28) характеризуются значительно меньшей частотой GLO1*1, чем народы Средней Азии (q = 0.39), Кавказа (q = 0.40), Урала (q = 0.41) и Восточной Европы (q = 0.42) [Балановская, 1998]. АРЕАЛ РУССКОГО ГЕНОФОНДА В русских популяциях (К = 18) частота аллеля GLO1*1 (рис. 5.2.12.) варьирует в довольно широких пределах 0.27 < q < 0.48 со средней q = 0.37. При этом GLO1 характеризуется несколько сниженной межпопуляционной изменчивостью (GST= 1.04) по сравнению с селективно-нейтральным уровнем русского генофонда (GST = 1.36). Генетический рельеф GLO1*1 в русском ареале крайне любопытен: максимальные значения частоты GL01*1 наблюдаются в центре ареала, а на периферии ареала они снижаются. Причём минимумы (0.27-0.30) обнаруживаются как в южной части ареала (включая часть волго-окского междуречья), так и в самой северной части (Архангельская область). ИТОГ Однако, исходя из данных о распределении аллеля GLO1*1 в коренном населении Европы, можно было прогнозировать совершенно иную картину для русского генофонда: увеличения частоты аллеля на юге (влияние южных европеоидов) и на севере, поскольку в циркумполярных финно- и самодийскоязычных популяциях частота GLO1*1 достигает значительных величин (например, у карел q = 0.44, у тундровых ненцев q = 0.51). Наблюдаемый генетический рельеф прямо противоположен ожидаемому, выведенному из основных геногеографических закономерностей у окружающих народов. Остается вновь предположить, что структура генофонда русского народа определяется не столько волнами иноземных миграций, сколько основными течениями собственной истории. Корреляции карт локуса GLO1 с картами всех трёх главных компонент русского генофонда невелики и колеблются в пределах 0.3 < r < 0.4. §1.9 СИСТЕМА 6-ФОСФОГЛЮКОНАТ-ДЕГИДРОГЕНАЗЫ (6PGD)Фермент необходим для синтеза нуклеиновых кислот (участвует в превращении гексоз) и присутствует во многих тканях организма. Кроме основных аллелей (PGD*A и PGD*C) описан ряд редких аллелей, причём некоторые приводят к сниженной активности фермента. Относится к группе генетико-биохимических маркёров (ферменты эритроцитарной фракции крови). В МАСШТАБЕ ОЙКУМЕНЫ Во всем мире гетерозиготность локуса мала за счёт резкого преобладания аллеля PGD*A. Самые низкие частоты PGD*C характерны для коренного населения Америки и Европы (q < 0.03), в остальных частях света q>0.05 с максимумами в Африке (q = 0.09), Меланезии и Индостане (q = 0.07). Между регионами Северной Евразии различия недостоверны: в Сибири q = 0.06, на Кавказе q = 0.05, в Средней Азии q = 0.05, на Урале q = 0.04, в регионе Восточной Европы (без Приуралья и Северного Кавказа) q = 0.04. Предполагается, что несколько большая частота аллеля обычно наблюдается для более южных популяций. АРЕАЛ РУССКОГО ГЕНОФОНДА В 12 изученных русских популяциях частота PGD*C слабо варьирует от 0.02 до 0.06 со средней q = 0.04 и гетерозиготностью HS = 0.035 (табл. 5.2.1). Уровень межпопуляционной изменчивости (GST = 0.35) значительно ниже селективно-нейтрального уровня (GST = 1.36), что может указывать на мощное действие стабилизирующего отбора. На карте (рис. 5.2.13) обращает на себя внимание ядро повышенных частот PGD*C в центральной и юго-восточной частях ареала. Однако оно в основном связано с интерполяцией на юго-восточные районы и базируется на данных о двух отнюдь не южных популяциях - Вологодской (q = 0.06, n = 50 чел.) и Московской (q = 0.06, n = 99 чел.) областей. Лишь дальнейшее изучение русского генофонда может проверить правильность представленного на карте генетического рельефа - слишком мало изученных популяций и слишком малы выборки для сравнительного изучения низко полиморфного гена. Поэтому эту карту можно оценивать лишь как один из вариантов картографического прогноза, который пока не может служить предметом детального обсуждения. Тем не менее, карта распространения PGD*C содержит важные структурообразующие элементы русского генофонда: корреляция карты PGD*C с картой первой главной компоненты изменчивости классических маркёров велика: r = 0.67. 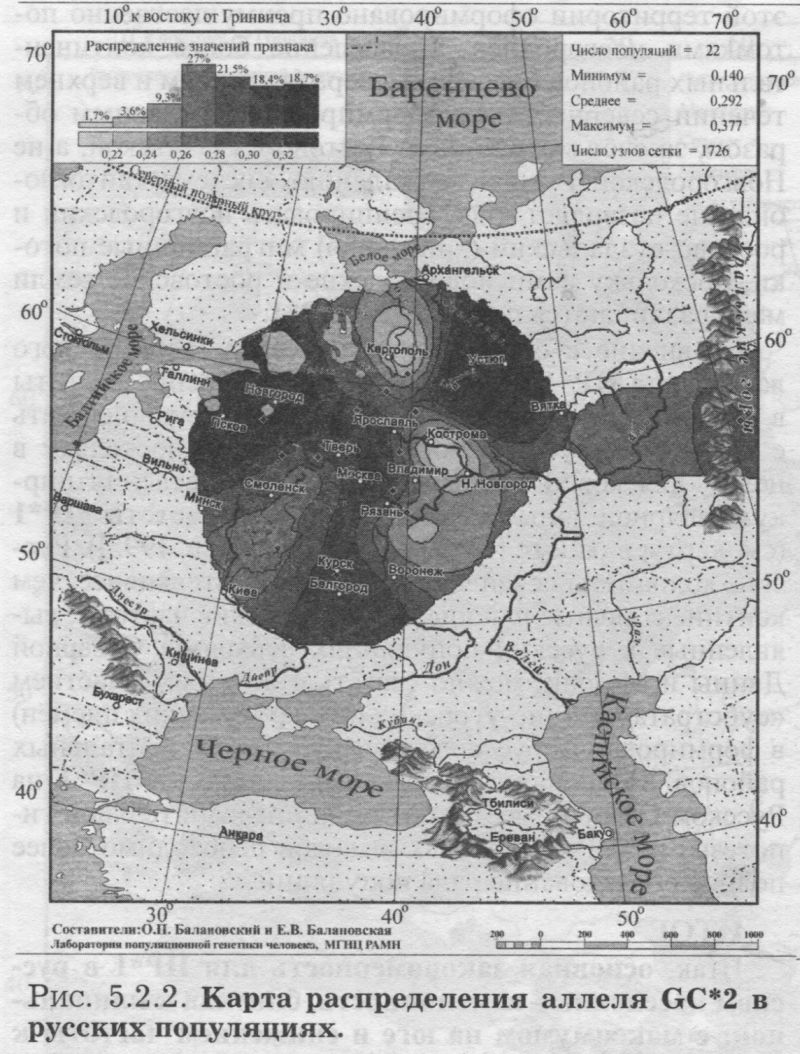
§2. Группы крови (MN, RH, АВО)Системы групп крови относятся к иммунологическим маркёрам - такова техника выявления этих маркёров. В самом простом случае это выглядит так. Если к эритроцитам, которые обладают так называемым «антигеном», мы добавим сыворотку крови другого человека, в которой есть соответствующие именно этому антигену антитела, то произойдет «слипание» эритроцитов. Мы увидим его невооруженным глазом: вместо обычной капли крови мы увидим сгустки и сыворотку (примерно так, как если в кислое молоко добавить кипяток). Обычно так и открывали многие иммунологические маркёры - если вдруг при переливании крови одного человека другому происходила агглютинация (слипание) эритроцитов, что могло приводить к осложнениям и даже гибели человека. Предназначение иммунологических маркёров - защищать организм от проникновения «чужеродцев», распознавать их и обезоруживать. Поэтому антитела могут вырабатываться организмом только к «чужим» антигенам - тем, которых у самого человека нет. §2.1 СИСТЕМА ГРУПП КРОВИ MNСистема MN относится к кодоминантным локусам: все три генотипа ММ, MN, NN проявляются фенотипически различно, что для иммунологических маркёров - редкость. Локус MN находится на 4 хромосоме. Тесно сцеплен с локусом Ss, образуя единую систему MNSs. Поэтому при совместном анализе обоих диаллельных маркёров выявляются четыре различных гаплотипа (MS, Ms, NS, Ns). Но поскольку по системе Ss и гаплотипам MNSs изучено меньшее число популяций, мы рассмотрим геногеографию лишь локуса MN. Между аллелями М и N нет доминирования одного над другим, поэтому гетерозиготу легко MN различить - она будет реагировать с обоими антителами - анти-М и анти-N. Различия между аллелями М и N связаны с аминокислотными заменами полипептидных цепей антигенной молекулы в позициях от 1 до 5. Антигены М и N обнаружены не только у человека, но и некоторых беспозвоночных. Выявлено присутствие антигенов М и N не только в крови, но и других тканях человека. Известны случаи осложнений после переливания крови, вызванных антителами анти-М. Выявлены некоторые ассоциации групп крови MN с сахарным диабетом, пневмонией, атеросклерозом, остеохондрозом. Однако физиологическая роль антигенов MN неизвестна. Относится к группе иммунологических генных маркёров. В МАСШТАБЕ ОЙКУМЕНЫ Гетерозиготность локуса MN достигает максимальной планки, возможной для двуаллельных локусов, поскольку в среднем для ойкумены частоты обоих аллелей близки к 0.5: среднемировая частота аллеля MN*M составляет q = 0.54. Причём равновесие между обоими аллелями поддерживается во всех регионах Старого Света: в Европе средняя частота аллеля MN*M q = 0.59, в Северной Евразии q = 0.57, в Индостане q = 0.66, в Восточной Азии q = 0.66, в Африке (южнее Сахары) q = 0.55, в Океании q = 0.45. Но, тем не менее, мировой размах частот максимально велик (от 0 до 1) за счёт своеобразия остальных континентов. Аллель MN*M редок в Новой Гвинее и Австралии, где его средняя частота равна q = 0.30, а в Меланезии - лишь q = 0.15. В коренном населении Америки, напротив, аллель MN*M преобладает, достигая средней величины q = 0.72. ЛАНДШАФТ ЕВРАЗИИ В пределах региона Северной Евразии, среди её локальных популяций, размах изменчивости аллеля MN*M также велик - от 0.17 до 0.90! Весь диапазон изменчивости приходится на популяции Сибирского региона, где величина межпопуляционных различий достигает чрезвычайно высокого уровня GST = 8.30. Причём в Сибири велики не столько различия между локальными популяциями в пределах этноса (то есть гетерогенность внутри этноса): по локусу MN они (GST = 2.09) даже меньше, чем в среднем для сибирских этносов (GST = 3.85, см. раздел 5.3.). Основная часть межпопуляционной изменчивости локуса MN (3/4) приходится на различия между этносами Сибири (GST = 6.21). Средние этнические частоты варьируют в необычайно широких пределах от q = 0.28 до q = 0.76 (размах межэтнической изменчивости Δ = 0.48). То есть этнические частоты у народов Сибири варьируют почти в тех же пределах, что и различия между локальными популяциями всей Северной Евразии (как говорилось, от 0.17 до 0.90). Причём не один, а целый ряд этносов (алеуты, долганы, негидальцы, ульчи, шорцы, якуты) характеризуется очень высокой средней частотой около q = 0.70. В то же время у другого ряда сибирских народов средние частоты едва достигают отметки q = 0.40 (нганасаны, ненцы, чукчи, чулымцы, энцы). При этом средняя частота для сибирского региона находится всё на том же уровне мировых стандартов: q = 0.56. На примере Сибири мы видим, что хотя среднерегиональные частоты достаточно постоянны в Старом Свете, внутри региона может наблюдаться значительный разброс частоты у разных народов, живущих в этом регионе. Среднерегиональная частота MN*M практически та же и в Уральском регионе: q = 0.52. И размах межпопуляционных различий MN также велик для населения Урала: GST = 4.2. Причём вновь, как и в Сибири, гетерогенность самих народов не велика: на Урале (GST = 1.04) она_по локусу MN меньше, чем в среднем для региона (GST =1.42). Как и в Сибири, основная часть (вновь %) изменчивости приходится на различия между коренными народами Урала GST = 3.20. Этнические частоты на Урале варьируют почти в столь же большом диапазоне, как и в Сибири - от q = 0.33 у ненцев до q = 0.68 у башкир (размах межэтнической изменчивости Δ = 0.35). В Средней Азии диапазон изменчивости этнических частот, напротив, чрезвычайно узок (от q = 0.56 до q = 0.62, то есть Δ = 0.06), а средняя региональная частота (q = 0.59) опять близка к «стандарту» Старого Света. Из всех регионов Северной Евразии только на Кавказе наблюдается некоторое повышение средней частоты (q = 0.68), а этнические частоты слабо варьируют от q = 0.59 у азербайджанцев до q = 0.76 у абазин (то есть размах межэтнической изменчивости Δ = 0.16). ГЕНОФОНД ВОСТОЧНОЙ ЕВРОПЫ Размах изменчивости частот MN*M среди народов Восточной Европы столь же невелик, как и на Кавказе: от q = 0.54 у литовцев до q = 0.70 у саамов (Δ = 0.16), причём практически у всех остальных народов Восточной Европы частоты колеблются около q = 0.60. Соответственно, средняя частота MN*M в Восточной Европе равна q = 0.62. Межпопуляционные различия по локусу MN (табл. 5.2.2.) очень малы (GST = 0.99) по восточноевропейским стандартам, где средний уровень различий между всеми популяциями Восточной Европы GST = 2.25 (см. подробно главу 8). Эта величина включает в себя как различия между популяциями в пределах народа (гетерогенность этноса), так и межэтническую изменчивость (различия между этносами), которая по локусу MN вообще неощутима: GST = 0.11 при средней по Восточной Европе GST = 0.82. Но и рассматривая только внутриэтнический уровень, мы видим сниженную изменчивость локуса MN: гетерогенность этноса по локусу MN (GST = 0.87) значительно меньше, чем селективно-нейтральная оценка гетерогенности «среднего» этноса восточноевропейского региона (различия между популяциями GST = 1.44). Эти сравнения, показывающие сниженную изменчивость локуса MN в Восточной Европе, суммированы в таблице 5.2.2. 
Изумляет и близость этнических частот восточных славян (причём оценки надёжны - каждый народ изучен по большому числу популяций): у русских q = 0.59, у белорусов q = 0.59, у украинцев q = 0.61. То есть при рассмотрении в целом восточноевропейской изменчивости MN бросается в глаза столь безликая межпопуляционная изменчивость, которая предполагает, что локус будет мало информативен для анализа генофонда. АРЕАЛ РУССКОГО ГЕНОФОНДА Напомним, что картографируемая территория включает русские популяции, расположенные от 43° до 70° с.ш. и от 24° до 60° в.д. (см. раздел 5.1). Поэтому в анализ попадает и часть территорий г. Прибалтики, Белоруссии, Украины. Однако лишь русское население этих территорий включено в анализ русского генофонда! В 32 изученных русских популяциях частота MN*M варьирует от 0.37 до 0.72 (табл. 5.2.1). Различия между русскими популяциями (GST = 1.19) чуть_ ниже селективно-нейтрального уровня (GST = 1.36). При этом гетерогенность русского народа равна общей изменчивости всех популяций Восточной 60° Европы по этому локусу (как мы сказали, для Восточной Европы по локусу MN GST = 1.20). То есть в восточноевропейском масштабе изменчивость русских популяций по этому гену велика и достигает размаха изменчивости всех популяций региона. Генетический рельеф аллеля MN*M в русских популяциях приведён на рис. 5.2.14. Рельеф карты задаётся как ядрами с высокой частотой гена, разместившимися и на самом юго-востоке, и на северо-западе ареала, так и областью низких значений на Русском Севере. Эти географически полярно расположенные элементы при почти полной аморфности остальных территорий и создают корреляцию признака с широтой r = 0.44 (табл. 5.2.1). Таким образом, мы оказываемся перед фактом, что при полной «безликости» и неинформативности средних частот MN*M, карта выявляет четкий тренд изменчивости в локальных русских популяциях. Присмотримся к нему. Ядра с высокими частотами MN*M на юго-востоке и северо-западе соединены «нагорьем» повышенных значений частоты. На Верхней Волге (Тверские популяции) это нагорье вырастает почти до уровня северо-западного ядра, формируя третье ядро повышенных значений. Отметим, что пики самых высоких частот аллеля М находятся за пределами «исконного» русского ареала (русские восточной Эстонии и восточной Украины). А русские популяции в «исконном» ареале удивительно похожи друг на друга и лежат в пределах двух средних интервалов (от 0.58 до 0.61 и от 0.61 до 0.64). Эти два интервала занимают две трети (65%) всей территории, образуя выраженное одновершинное распределение {рис. 5.2.14). Сразу подчеркнём, что «ядра» высоких значений нельзя однозначно вывести из частот у окружающих народов. Частоты русских популяций северозападного ядра выше, чем в окружающих популяциях эстонцев (q = 62, оценка получена по большому числу - 15 популяций эстонцев). Частоты в юго-восточном ядре также выше, чем у восточных соседей русского народа - в популяциях калмыков (q = 0.64), татар (q = 0.54), мордвы (q = 0.57), удмуртов (q = 0.59), марийцев (q = 0.56). Оказывается, что частоты MN*M в окружающих этносах находятся в пределах всё тех же двух средних интервалов (от 0.58 до 0.64) или даже ниже их, в то время как в окраинных «ядрах» русских популяций значения (q = 0.72) заметно превосходят значения средних интервалов (от 0.58 до 0.64). Вторым структурообразующим элементом является область низких частот на севере8. Она задаётся шестью популяциями Архангельской области в бассейне Мезени, изученными А. А. Ревазовым и его сотрудниками. Все шесть популяций попадают в интервал значений q < 0.55. В одной из популяций частота опускается до q = 0.37, а в остальных колеблется около 0.5. Выборки велики - в среднем по 140 человек из популяции. Поэтому снижение частоты MN*M на севере Архангельской области можно считать твёрдо установленным фактом. ГИПОТЕЗЫ Достойное объяснение станет возможным тогда, когда и иные популяции Русского Севера будут детально изучены по MN. Однако и теперь можно высказать предположение, что такое снижение частоты на севере русского ареала связано с влиянием дославянского населения, включенного в русский генофонд в процессе колонизации северных территорий. Эта гипотеза имеет основания в низкой частоте у северовосточных уралоязычных групп. Градиент частоты у них аналогичен русским популяциям - частота падает к северу (у коми-пермяков q = 0.54, у коми-зырян q = 0.50, у ненцев q = 0.33). Но речь может идти о влиянии именно восточных групп финно-угров, поскольку у западных финноязычных групп частоты MN*M выше, чем у северных русских: у карел q = 0.62, у вепсов q = 0.64, у эстонцев q = 0.62, у саамов q = 0.70. В порядке планирования будущих работ можно высказать ещё одну, ничем не обоснованную, но заманчивую гипотезу. Она касается того «коридора» сниженных частот MN*M, который тянется через весь русский ареал с севера на юг примерно вдоль 32 меридиана. Велик соблазн объяснить его едва сохранившимся следом пути из «варяг в греки», протянувшимся через озера, реки и волоки от Ладоги до Черного моря. Проверить эту гипотезу просто - надо лишь пройти с экспедиционным обследованием этим же путем, что ходили наши предки. Корреляция карт аллелей локуса MN выявлена лишь с картами третьей главной компоненты русского генофонда: r = 0.5. §2.2 СИСТЕМА ГРУПП КРОВИ РЕЗУС(RH)Система групп крови Резус всем известна по личному знакомству - её определяют, когда возникает риск иммунологического конфликта: матери и плода, или же при переливании крови. Три тесно сцепленных локуса расположены на 1 хромосоме в порядке D-C-E, образуя единую систему Rhesus (RH), и при анализе всех трёх локусов можно выявить 18 фенотипов, за которыми скрываются 8 гаплотипов. Но поскольку по гаплотипам D-C-E изучено меньшее число популяций, чем только по локусу D, то мы и рассмотрим геногеографию лишь этого локуса. В популяционно-генетических исследованиях он представлен как двуаллельная система (RH*D и RH*d). И именно с локусом RH-D связана основная угроза возникновения гемолитической болезни новорожденных, хотя другие два локуса RH-C и RH-E также могут быть причиной иммунологического конфликта между плодом и матерью. Антигены образуются у плода очень рано - уже в возрасте 8 недель. Небольшой размер иммуноглобулинов, несущих антитела анти RH, позволяет проникать им через плацентарный барьер, и иммунизация материнского организма приводит к гемолитической болезни новорожденного. Гомозиготы dd по рецессивному аллелю RH*d (то есть обладатели двух рецессивных аллелей RH*d) всем известны как «резус-отрицательные»; а гомозиготы DD и гетерозиготы Dd (то есть носители хотя бы одного доминантного аллеля) фенотипически неразличимы и известны нам как «резус-положительные». Поясним, что именно из-за доминирования RH*D над RH*d (d внешне никак не проявляется в присутствии D, т. е. в гетерозиготе Dd) и возникают недоумения: почему у «резус-положительных» супругов рождаются «резус-отрицательные» дети? Это происходит, когда оба супруга гетерозиготны (Dd): ведь каждый из них является носителем рецессивного аллеля d, не проявляющегося «внешне», фенотипически. Из-за доминирования D над d оказываются возможны и разные исходы в браках «резус-положительных» и «резус-отрицательных» супругов. В этом случае все зависит от генотипа резус-положительного супруга. Все дети будут «резус-положительными», если его генотип окажется гомозиготой DD. Но если он гетерозиготен (Dd), дети будут «резус-отрицательные» с вероятностью 0.5. Показана связь системы RH со многими заболеваниями - язвой желудка, остеоартрозом, остеохондрозом, пневмонией, корью, краснухой, а также связь с гормональным статусом. Относится к группе иммунологических генных маркёров. В МАСШТАБЕ ОЙКУМЕНЫ Географическая изменчивость аллеля RH*d, ответственного за «резус-отрицательность», общеизвестна: максимум на западе Евразийского континента у басков Испании и Франции (q = 0.74) и постепенное снижение частоты во всех остальных направлениях. Частоты аллеля RH*d в регионах мира приведены в таблице 5.2.3. Даже из таблицы видно, что мир как бы распадается по RH*d на две неравные части. С одной стороны запад Евразии с присоединившимися к нему Африкой, Уралом и Средней Азией. В этих регионах частоты RH*d заметны. С другой стороны - мир без RH*d (частоты RH*d близки к нулю). Это восток Евразии с обеими Америками, Австралией, Меланезией, Океанией. Северная Евразия занимает особое положение, охватывая оба мира - с RH*d и без него. Поэтому в таблице приведены частоты в регионах мира, а для Северной Евразии - ещё и для отдельных субрегионов. 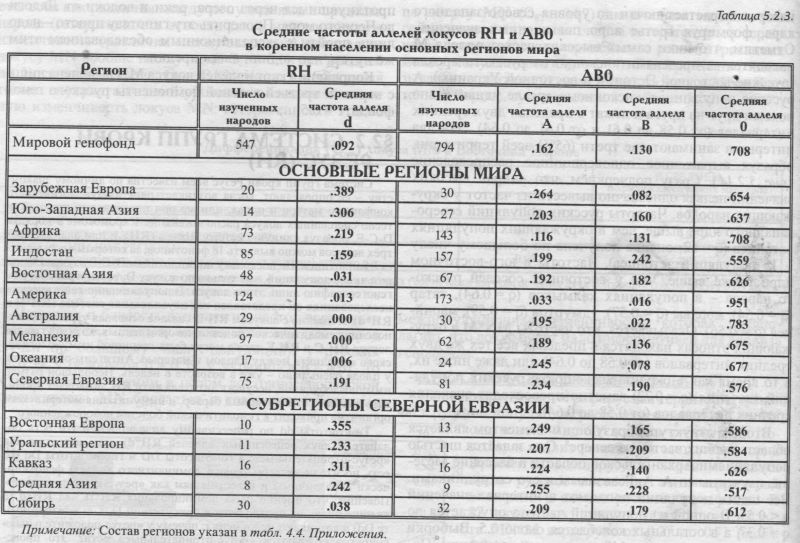
ЛАНДШАФТ ЕВРАЗИИ (рис. 5.2.15.) Прежде всего, поясним, что создание карт евразийских трендов RH*d и АВ0 не было самоцелью. При формировании Банка данных «GENEPOOL» нас интересовал генофонд Северной Евразии на фоне его соседей. Поэтому информация для народов бывшего СССР собиралась максимально тщательно, а прочие регионы характеризовались, конечно, не столь полно. Хотя Европа и покрыта густой сетью изученных популяций, но часть Центральной, Южной и Передней Азии (и тем более Африки) представлены в Банке данных, по нашей лености, лишь отдельными народами. Это не мешает прослеживанию основных трансевразийских трендов. Но хотелось предостеречь читателя от чрезмерного доверия к локальным закономерностям карт RH*d и АВ0 Евразии. Впрочем, это относится к любым картам и таблицам - несовершенство наших знаний всеобъемлюще. Поэтому мы всегда привлекаем внимание читателя к наиболее устойчивым «каркасам» генофонда, его основным закономерностям, а мимолетное и эфемерное стараемся оставить за рамками портрета. Для решения этой же задачи мы приводим карты Евразии слегка «сглаженные» (фоновые, трендовые - см. специально посвященную этим технологиям статью [Балановская, Нурбаев, 1995] и Приложение). Это позволяет избавиться от случайных флуктуаций в частоте аллелей и привести на «сглаженной» карте наиболее устойчивые значения, достойные внимания и доверия читателя. Генетический ландшафт RH*d в Евразии не требует специальных пояснений - настолько он изысканно прост (рис. 5.2.15). Максимумы частот RH*d сосредоточены только в Западной Европе. При движении в остальных направлениях частота RH*d плавно снижается. Несомненен основной тренд: неуклонное падение частоты RH*d с запада на восток Евразии. Причём изолинии, делящие диапазон изменчивости на равные интервалы, почти вторят направлению меридианов (рис. 5.2.15.) Границей служит 90й меридиан - коренное население к востоку от него всё «резус-положительно». Восточная Европа - предмет нашего главного интереса - являет собой переходную зону, хотя в целом принадлежит к «европейскому» резус-отрицательному миру. СЕВЕРНАЯ ЕВРАЗИЯ (бывший СССР) Среди локальных популяций Северной Евразии размах изменчивости аллеля RH*d хотя и уступает мировому, но очень велик - от 0.00 до 0.54! Причём столь широкий диапазон изменчивости (от 0 до 0.5) среди локальных популяций характерен и для субрегионов Северной Евразии: и Восточной Европы, и Урала, и Средней Азии. Лишь на Кавказе минимальная частота опускается не до 0, а до q = 0.16. Зато в коренном населении Сибири и размах изменчивости мал, и сам аллель RH*d встречается очень редко - в среднем q = 0.04. Среди сибирских народов RH*d достигает заметных значений лишь у народов Алтае-Саянского нагорья (у шорцев q = 0.17, у тофаларов q = 0.14, у хакасов q = 0.11). На Урале сниженная средняя частота гена RH*d (q = 0.23) связана с популяциями Зауралья и самого севера Урала (у манси q = 0.16, у хантов q = 0.00, у тундровых ненцев q = 0.10). Наиболее близкие восточные соседи русского народа - популяции Приуралья и Южного Урала - по частотам RH*d приближаются к народам восточно-европейского региона: у коми-зырян q = 0.30, коми-пермяков q = 0.26, мари q = 0.37, мордвы q = 0.36, чувашей q = 0.29, татар q = 0.30, башкир q = 0.27, и только у удмуртов q = 0.17 (оценка получена по 13 популяциям). Межпопуляционная изменчивость RH*d на Урале необычайно велика: GST = 7.9. ВОСТОЧНАЯ ЕВРОПА В Восточной Европе средние различия по RH*d между всеми её локальными популяциями в два раза меньше, чем на Урале, и равны GST> = 3.6. Первая составляющая этой величины - гетерогенность «среднего» восточно-европейского народа - по RH*d оценивается как GST = 0.87, что несколько ниже селективно-нейтральной оценки GST = 1.44 (табл. 5.2.4). Однако вторая составляющая - различия между народами -_по локусу RH*d достигает очень большого размаха (GST = 2.75), который_в три раза больше восточноевропейского стандарта (GST = 0.82). Эти сравнения суммированы в таблице 5.2.4. 
В диапазон этнических частот от q = 0.33 у эстонцев и вепсов до q = 0.44 у соседних карел «уложились» все народы Восточной Европы. Лишь у саамов (лопарей), своеобразие которых проявляется практически по любым маркёрам, средняя частота RH*d очень низка: q = 0.14. Без учета саамов, средняя частота RH*d в Восточной Европе составляет q = 0.38. Такая же средняя частота свойственна и русскому этносу: q = 0.38 (оценка получена только по тем 96 русским популяциям, которые участвовали в построении карты на рис. 5.2.16). Между восточными славянами, где каждый народ изучен по RH*d очень подробно (по 70-100 популяциям), различия средних частот невелики и варьируют около «русских» частот: у белорусов q = 0.41, у украинцев q = 0.34. Характер изменчивости RH*d в Восточной Европе лучше описать с помощью корреляционных карт, показывающих связь частоты гена с широтой и долготой в каждой точке карты (см. карты [Балановская, Нурбаев, 1994а,б; 1995]). Для основной части Восточной Европы9 характерна чрезвычайно высокая корреляция с долготой, демонстрирующая падение частоты гена RH*d в направлении «запад-восток» (rMIN = - 0.80). Именно такой тренд и ожидается, исходя из простой изменчивости аллеля в Евразии в целом (рис. 5.2.15). Действительно, высокая отрицательная корреляция с долготой означает, что большему значению географической долготы соответствует меньшая частота аллеля. Поскольку для Восточной Европы значения долготы растут с запада на восток (отсчитываясь, как известно, от Гринвичского нулевого меридиана), мы получаем снижение частоты аллеля RH*d к востоку. Этот пример показывает, как, пользуясь одними лишь корреляционными картами, можно извлекать подробную информацию о географии аллеля - в данном случае мировой тренд падения частоты аллеля к востоку. Однако при переходе от мира на уровень региона, регион обнаруживает и свои собственные закономерности, нарушающие стройный ход мировых закономерностей. Так, северо-западная зона Восточной Европы обнаруживает «обратное» направление связи с долготой (rMAX= + 0.60). Что же касается корреляции с широтой, то средний коэффициент корреляции приближается к нулю (r = 0.01). Однако нулевые корреляции с широтой (- 0.10 < r < +0.10) занимают лишь 11% всей площади карты. Выделяются две области: высокой отрицательной связи с широтой (rMIN = - 0.86) на севере и почти столь же высокой положительной связи с широтой (rMAX = + 0.56) на юге Восточной Европы. Корреляционные карты говорят о двух важных явлениях. Во-первых, что средний коэффициент корреляции (то есть обычный, который мы рассчитываем без карт) может ввести в заблуждение. Если связь между признаками мощная, но существуют две равных по площади области с противоположной по знаку (до r= +1 и r = -1) связью, то средний коэффициент становится равным нулю, и мы можем решить, что связи просто нет! Во-вторых, переходя от глобального рассмотрения к составляющим регионам (все более маленьким «матрёшкам») мы можем обнаруживать исчезновение «глобальных» закономерностей и появление своих собственных, региональных. АРЕАЛ РУССКОГО ГЕНОФОНДА Именно это мы и обнаруживаем для русского генофонда. На карте русского народа (рис. 5.2.16.) мы не видим даже намёка на общий трансевразийский тренд «запад-восток» (рис. 5.2.15). Более того, мы не видим вообще никаких четких закономерностей в ареале русского генофонда! «Русская» карта RH*d подобна «лоскутному одеялу»: на ней области высоких значений мирно соседствуют с областями низких значений RH*d. Если же выделить из всей картографируемой территории10 тот основной «исконный» ареал, в котором складывался физический облик русского народа11, то в нём можно разглядеть не очень четкий, но своеобразный рельеф. В центре ареала находятся два пика (р>0.42) и «нагорья» высоких значений, от которых во всех направлениях частота гена 7() RH*d снижается. На периферии ареала мы видим либо новые повышения частоты (на юге и востоке), либо снижения (на севере, западе и юго-востоке). Напомним, мы ожидали для локуса MN, что (из-за его безликой изменчивости в Восточной Европе) он будет мало информативен для анализа русского генофонда (подраздел §2.1.), но обнаружили в русском ареале географический тренд с широтной изменчивостью! Для RH*d картина обратная - ожидая, исходя из глобального тренда, простую закономерность «запад-восток», мы сталкиваемся со столь причудливым генетическим рельефом, который даже трудно интерпретировать! Карты показывают, что географическое распространение гена не укладывается в простую схему, предполагающую, что континентальный тренд выложен, как из «кирпичиков», из внутрирегиональных и внутриэтнических трендов, и каждый из этносов отражает ту часть всеобщего тренда, которая приходится на его территорию. Обычно все оказывается сложнее и любопытнее. Например, ожидаемый для RH*d тренд (убывание частоты гена в направлении «запад-восток») ярко проявляется лишь при рассмотрении евразийского континента в целом (рис.5.2.15). Но с изменением масштаба исследования, при последовательном переходе к более частным субрегионам и далее к этносам (см. карты [Балановская, Нурбаев, 1994а,б; 1995]), за деревьями исчезает лес - континентальный тренд многократно перекрыт более частными событиями истории этих генофондов. Возвращаясь к русскому генофонду, отметим, что в 96 изученных русских популяциях частота RH*d варьирует от 0.26 до 0.52 (табл. 5.2.1). Но при этом гетерогенность RH*d (GST = 0.45) намного ниже селективно-нейтрального уровня (GST = 1.36) гетерогенности русских популяций. Столь резкое снижение изменчивости может указывать на действие стабилизирующего отбора на локус RH-D в русских популяциях. При этом изменчивость русского народа по RH*d (GST = 0.45) меньше, чем многих других восточно-европейских народов: она в два раза слабее усредненной гетерогенности народов Восточной Европы по этому локусу (GST = 0.90, табл. 5.2.4). Напомним, что на межэтническом уровне различия между народами Восточной Европы по RH*d ещё значительней: GST = 2.7. В целом, это означает, что трансевразийский тренд RH*d «запад-восток» проявляется в Восточной Европе лишь за счёт иных народов и, скорее всего, на уровне межэтнических различий, но не имеет отношения к русскому народу и не проявляется в гетерогенности русских популяций. Локус RH*d вносит весомый вклад во второй главный сценарий архитектоники русского генофонда: r = 0.73. §2.3 СИСТЕМА ГРУПП КРОВИ АВ0Локус АВ0 является самым известным из всех генов человека - все знают его как просто «группы крови» - I(0), II(A), III(B), IV(AB). Группы крови АВ0. как и Резус, определяют возникновение риска иммунологического конфликта при переливании крови. Благодаря их широкому применению в медицинской практике и, следовательно, обилию данных по разным популяциям, в течение долгого времени эти маркёры являлись излюбленными, а то и просто единственными, маркёрами в арсенале популяционных генетиков. И сейчас группы крови АВ0 и Резус являются абсолютными лидерами по обилию популяционных данных. Пока можно лишь мечтать, что в будущем какой-либо маркёр будет, так же как АВ0, изучен в 182 русских популяциях и в 1230 популяциях Северной Евразии! Число аллелей в локусе АВО велико (особенно выявляемых методами молекулярной генетики), но общеизвестные «группы крови» определяются только тремя аллелями - А, В и 0. Аллели А и В кодоминантны («равноправны») и поэтому гетерозигота АВ проявляется фенотипически - как «четвертая» группа крови IV(AB). Аллель 0 рецессивен («подчинён») по отношению и к А, и к В. поэтому фенотипически мы обнаруживаем лишь его гомозиготу, обозначая её как «первую» группу крови I(0). Из-за такого «подчинения» гетерозигота А0 фенотипически неотличима от гомозиготы АА, и потому вместе они образуют «вторую» группу крови I(A). Точно также оба генотипа В0 и ВВ внешне проявляются как «третья» группа крови III(B). Знание такого «соподчинения» аллелей позволяет ожидать, что в браке супругов с I группой крови все дети будут им подобны. В браке супругов с IV группой крови - дети могут быть любой группы крови, кроме I. А в браке супругов с II и III группами возможны все варианты - от I до IV групп крови (если оба супруга гетерозиготны, т. е. А0 и В0). Антигены образуются у плода очень рано - уже в возрасте 5 недель. Поэтому при несовместимости матери и плода по АВ0. может возникнуть иммунологический конфликт, который приводит не столько к гемолитической болезни новорожденного, сколько к спонтанным абортам. Но они обычно происходят столь рано, что о них и не подозревают. Если антитела из сыворотки крови матери проникают через плацентарный барьер в кровь ребенка, то они могут вызвать агглютинацию (слипание) его эритроцитов, что приводит к гибели плода на ранних стадиях эмбриогенеза. Такой отбор на ранних стадиях развития плода можно обнаружить лишь статистически - в нехватке детей с определенными фенотипами среди потомства в несовместимых по АВ0 семьях. В более поздние сроки беременности внутриутробная гибель плода приводит к спонтанным выкидышам. Показана связь групп крови АВ0 со многими заболеваниями. Вирусный гепатит, грипп, краснуха, скарлатина, язва желудка, язва кишечника, многие виды рака, болезни опорно-двигательного аппарата, заболеваний дыхательных путей и сердечно-сосудистой системы - вот далеко не полный перечень болезней, устойчивость или же подверженность к которым коррелирует с группами крови АВ0. Географическую неравномерность в мировом распределении групп крови АВ0 часто связывают с результатами эпидемий чумы, оспы и холеры: обладатели группы крови I(0) более восприимчивы к холере и чуме, чем обладатели группы крови II(A). Причину этого видят в том, что палочка чумы содержит антиген, подобный антигену 0, а вирус оспы - подобный антигену А. Поэтому, например, индивиды с группой крови I(0), имеющие антиген 0. могут оказаться менее устойчивыми к чуме, потому что им сложнее «опознать» антиген возбудителя болезни как чужеродный и они могут реагировать на него слабее и несвоевременно. Именно с этим феноменом принято связывать низкие частоты группы крови I(0) в регионах, где свирепствовали эпидемии чумы: Северная Африка, Индия, Монголия. Относится к группе иммунологических генных маркёров. В МАСШТАБЕ ОЙКУМЕНЫ Описание изменчивости аллелей АВ0*А, АВ0*В, АВ0*0 в масштабе ойкумены кочует из учебника в учебник. Поэтому мы ограничимся приведенной выше таблицей с рассчитанными нами средними частотами для всех регионов мира (табл. 5.2.3.) и картами распространения аллелей АВ0 для населения Евразии (рис. 5.2.17.-5.2.19). За пределами Старого Света (табл. 5.2.3.) больше всего поражает резкий сдвиг частот АВ0 в коренном населении Америки - повсеместно распространен аллель AB0*0 (q = 0.95), а частоты А и В равно близки к нулю. В коренном населении Австралии также очень высока частота AB0*0 (q = 0.78), но при этом аллель АВ0*А (q = 0.20) заметно преобладает над AB0*B (q = 0.02). В населении Евразии частоты AB0*0 (q = 0.61) ниже, но при этом ни в одном из евразийских регионов они не опускаются за отметку q = 0.5 (табл. 5.2.3.). Аллели А и В в разных регионах Евразии уступают друг другу второе место. В среднем для Евразии частота АВ0*А (q = 0.21) несколько выше, чем частота AB0*B (q = 0.18). В регионах Евразии оба аллеля достигают высоких частот в Индостане, где наиболее снижена частота мажорного нулевого аллеля. Описание евразийского распределения групп крови АВ0 несколько выходит за рамки русского генофонда. Поэтому будем предельно кратки - карты говорят сами за себя. Приводя эти карты, мы вновь делаем исключение, как и для групп крови Резус. И причина этому все та же - карты групп крови АВ0 вызывают всеобщее любопытство. Однако такое исключение помогает решить и непосредственную задачу книги - позволяет на примере наиболее изученных и надежных маркёров рассмотреть, как русский генофонд выглядит «из космоса» и как он вписывается в евразийские ландшафты. ЕВРАЗИЙСКИЙ ЛАНДШАФТ АВ0*А (рис. 5.2.17.) Он не столь элегантен, как ландшафт RH*d (рис. 5.2.15.), но все же позволяет, генерализуя картину, выделить такой же общий тренд: снижение частоты АВ0*А с запада на восток Евразии. На западе Евразии преобладают нагорья высоких значений АВ0*А, достигая максимальных значений и на самом севере Европы (Фенноскандия), и на самом её юге (Иберийский полуостров). Возвышенность генетического рельефа в Восточной Европе простирается на Украину и юго-запад России. Однако основной ареал Восточной Европы, включая Прибалтику, центр Русской равнины и особенно Русский Север отличаются от Западной Европы более низкими частотами АВ0*А. За пределами Европы значения выше средних частот проходят поясом через Переднюю и Среднюю Азию и далее на восток, вновь возрастая в Японии. К северу и к югу от этого пояса мы видим низкие значения АВ0*А. Впрочем, эта закономерность не всеобщая: например, в Сибири диапазон изменчивости АВ0*А достигает евразийского размаха. ЕВРАЗИЙСКИЙ ЛАНДШАФТ АВ0*В (рис. 5.2.18.) Этот ландшафт более выразителен. В Европе сосредоточены минимумы частоты АВ0*В, плавно возрастающей при движении на восток. Второй «полюс» - высоких частот АВ0*В - охватывает огромные пространства Южной и Центральной Азии. Итак, основной тренд: повышение частоты АВ0*В с запада на юго-восток Евразии. В отличие от предыдущей карты, генетический ландшафт Восточной Европы по АВ0*В не отличается своеобразием и исполняет роль переходной зоны: на западе она выглядит вполне европейски, а на востоке встраивается в долготный пояс высоких частот. Этот пояс пересекает Евразию с юга на север и тянется от Индийского океана (с мировым максимумом в Индостане) к Северному Ледовитому океану (к тундровым ненцам). В Сибири диапазон изменчивости частот АВ0*В опять-таки достигает евразийского размаха. Но к югу от Сибири мы видим устойчивое повышение частоты АВ0*В, характерное почти для всей южной части Азии. ЕВРАЗИЙСКИЙ ЛАНДШАФТ АВ0*0 (рис. 5.2.19.). Ландшафт этого аллеля особенно хорош с методической точки зрения. Он показывает, что ареалы с одинаковыми значениями признака вовсе не говорят о родстве населения, там обитающего. Все зависит от географии, от взаимоположения этих ареалов, от их разделённое™ пространством и историко-географическими барьерами. «Положение объекта в пространстве географы считают таким же существенным признаком, как физические свойства и функции» [Родоман, 1990, с.37]. Первой огромной зоной высоких частот АВ0*0 является Сибирь с прилегающими к ней районами. Несмотря на значительные колебания частот, связанные с дрейфом генов в сибирских изолятах, Северная Азия представляет собой обширное нагорье высоких частот АВ0*0. Вторая зона максимумов расположена на западе Западной Европы. Эти два максимума разделяет обширнейшая низменность со значениями АВ0*0 ниже евразийской средней. Эта область простирается от Центральной и Северной Европы до Японии, захватывая Восточную Европу, Урал, юг Западной Сибири, Центральную Азию и почти всю Южную Азию. Поэтому, при обнаружении такого ландшафта - с максимумами, разделёнными пространством, историей формирования населения и областями иных значений - нам не приходит мысль о близком родстве населения Сибири и Западной Европы. Объяснение такого ландшафта обычно ищут в терминах естественного отбора. Для групп крови АВ0 отбор, как считается, связан с распространением инфекций (оспы, чумы) и других заболеваний, опосредованных демографией (плотностью населения) и внешней средой - в самом широком смысле этого понятия. Однако и эта гипотеза оставляет много недоумений. Возвращаясь к Восточной Европе, - она наиболее важна для понимания русского генофонда - отметим своеобразие её северных областей. В них расположен локальный максимум частоты АВ0*0, заметный даже в евразийском масштабе. Причём на исходной карте (напомним, что мы приводим слегка «сглаженные» карты, чтобы выявить устойчивые тренды) этот локальный максимум отделен от соседей как на западе, так и на востоке. СЕВЕРНАЯ ЕВРАЗИЯ На примере двух столь подробно изученных локусов - групп крови АВ0 и RH - можно убедиться, насколько различна бывает дифференциация одних и тех же регионов по разным маркёрам. Например, на Урале дифференциация популяций по RH*d (GST = 7.9) в пять раз выше, чем по АВ0 (GST = 1.5). В Восточной Европе - по RH*d (GST = 3.6) в два раза выше, чем по АВ0 (GST = 1.6). А на Кавказе показатели идентичны - GST = 2.4 и по RH*d, и по АВ0. Этот уровень межпопуляционных различий по АВО одинаково невелик на Урале (GST = 1.5) и в Восточной Европе (GST = 1.6), хотя в других регионах Северной Евразии он выше: на Кавказе GST = 2.4. в Средней Азии GST = 2.5, в Сибири GST = 5.5. Рассмотрим для краткости изменчивость лишь «ведущего» аллеля - АВ0*0, а особенности двух других аллелей читатель сможет сам проследить по картам и таблицам. В генофондах Северной Евразии наибольшая частота АВ0*0 обнаруживается на Кавказе (q = 0.63), причём такая же частота характерна и для Западной Европы (q = 0.65). У народов Восточной Европы и Урала частота АВ0*0 несколько ниже (q = 0.58). В обоих регионах частота АВ0*0 колеблется в широких пределах. В уральском регионе от q = 0.49 у тундровых ненцев до q = 0.64 у манси Зауралья. При этом тюркоязычные народы (башкиры q = 0.56, татары q = 0.57, чуваши q = 0.56) неотличимы от финно-угров (мари q = 0.56, мордва q = 0.57, удмурты q = 0.58, коми-пермяки q = 0.59, коми-зыряне q = 0.60) У народов Восточной Европы размах изменчивости вторит уральскому: от q = 0.47 у крымских татар до q = 0.63-0.64 у карел, вепсов и литовцев. ВОСТОЧНАЯ ЕВРОПА На межэтническом уровне, различия между восточнославянскими народами - русскими, украинцами, белорусами - невелики: 0.25 < q < 0.26 по АВ0*А; 0.14 На внутриэтническом уровне, гетерогенность русского народа по группам крови АВО намного ниже селективно-нейтрального (GST=1.36) уровня: для AB0*A G =0.88, для AB0*B GST=0.61, для АВ0*0 GST=0.76 (табл. 5.2.1). Это может указывать на действие стабилизирующего отбора на локус АВО. Гетерогенность не только русского, но и «среднего», «типичного» этноса восточноевропейского региона по АВО (GSt = 0.48) значительно - в три раза! - ниже селективно-нейтральной оценки (GSt=1.44, табл. 5.2.5.). Однако отсюда следует, что гетерогенность русского народа по АВО (Gst = 0.8) больше, чем гетерогенность «типичного» этноса Восточной Европы по этому локусу (Gst = 0.5). «РУССКИЙ» ЛАНДШАФТ АВ0*А (рис. 5.2.20.) В отличие от «русской» карты по RH*d, обнаруживается не «лоскутное одеяло», а четкий тренд. Ясно прослеживается падение частоты аллеля по направлению к северо-востоку: если в западных и южных русских областях частота достигает q = 0.30, то на северо-востоке практически нигде не превышает q = 0.22. Основная часть популяций лежит в пределах 0.15 < q < 0.35 (рис. 5.2.20). Несмотря на относительно небольшой размах изменчивости, всё множество популяций соответствует картине снижения частоты к востоку и северу. Удивительно, что лишь одна южная популяция (на нижней Волге) и одна северная (на Кольском полуострове) выбиваются из общего строя, а остальные 180 популяций следуют ему. Такое снижение частоты к востоку в русском ареале является частью общего тренда изменения частоты АВ0*А в Евразии в целом (рис. 5.2.17.) - падения частоты при движении от Западной Европы к Восточной и далее за Урал в Центральную Азию [Cavalli-Sforza et al., 1994]. 
«РУССКИЙ» ЛАНДШАФТ АВ0*0 (рис. 5.2.21.) Он является почти зеркальным отражением предыдущей карты - падение частоты с северо-востока (р>0.7) на юг и юго-запад (q < 0.5). Если на Русском Севере частота достигает 0.7, то в западных и южных русских областях снижается до 0.5. Основная часть популяций лежит в пределах 0.5 < q < 0.7. И вновь почти все русские популяции следуют рельефу плавного снижения частоты с севера к юго-западу, вновь подтверждая объективность такого тренда для русских популяций. Основное отличие карт этих двух аллелей (АВ0*А и АВ0*0) связано с северозападной частью ареала. По АВ0*А в ней наблюдается максимум высоких значений частоты - пик, сформированный Псковскими популяциями и завершающий гряду западных и юго-западных высоких значений. По аллелю АВ0*0 северо-запад тяготеет не к южным, а к северным зонам, являясь их закономерным продолжением. Повышение частоты АВ0*0 на юго-востоке лежит в области интерполяции и сопровождается сниженной (хотя и достоверной) надёжностью такого прогноза. Это сомнительное ядро (находящееся далеко за пределами «исконного» русского ареала) опирается лишь на три популяции со значениями частоты выше 0.6, в то время как для всего множества остальных русских популяций Поволжья характерны частоты в интервале 0.58 < q < 0.60. Выше мы привели средние частоты АВ0*0 для коренных народов Уральского региона - все они похожи друг на друга и лежат в узких пределах 0.56 < q < 0.58, достигая q = 0.60 лишь у коми-зырян на севере. У калмыков частота АВ0*0 также невысока: q = 0.57. У казахов q = 0.58 (это самая высокая частота из всех народов Средней Азии и Казахстана). Поэтому повышение частоты в трёх русских популяциях Поволжья, скорее всего, случайно: оно не соответствует частотам ни в соседних русских, ни в соседних иноэтничных популяциях. РУССКИЙ СЕВЕР (АВ0*0) Что же касается повышения частоты АВ0*0 на Русском Севере, то его нельзя объяснить влиянием народов Приуралья. Хотя у них мы обнаруживаем некоторое повышение частоты АВ0*0 к северу, но в северных русских популяциях частота гена АВ0*0 ещё выше и варьирует около 0.7. Коренные народы с высокими значениями АВ0*0 мы находим на северо-западе: и у карел, и у вепсов q = 0.64. В целом, опираясь на эти данные и историю колонизации Русского Севера, можно предположить, что повышение частоты АВ0*0 в северных русских популяциях связано с финно-угорским субстратом - теми племенами и народами, которые освоили эти территории задолго до славян. Причём можно предположить, что они были больше похожи на современных западных, чем на восточных финно-угров. Напомним, что по аллелю MN*M мы также обнаружили снижение частоты на Русском Севере, которое также объясняли влиянием финно-угорского субстрата (подраздел §2.1). Но по локусу MN вырисовывалась картина влияния не западных, а напротив, восточных групп финно-угров. Это показывает, сколь шатки любые выводы о генофонде, основанные на изменчивости лишь одного гена. Впрочем, нельзя исключить, что в генофонде Русского Севера присутствуют как западный, так и восточный финно-угорский субстрат. И разные гены зафиксировали в своем генетическом ландшафте только один из этих генных потоков. Поэтому описание устойчивой структуры русского генофонда мы даем в разделе 5.3, основываясь на обобщённом анализе всех классических маркёров. А предположениям, формулируемым при описании отдельных генов, отводим ранг лишь предварительных (и, как видим, порой противоречащих друг другу) гипотез. Однако рискнем высказать ещё одну гипотезу (см. также разделы 2.3. и 4.4). В генетическом ландшафте АВ0*0 на Русском Севере проявляется не только дославянский субстрат, но и потоки славянской колонизации. Исходя из общего генетического ландшафта, можно предположить, что для Новгородской Руси и её северных колоний были характерны более высокие значения АВ0*0. А «низовская» колонизация Ростово-Суздальской, а затем Московской Руси несла на север более низкие частоты АВ0*0, характерные для большинства летописных славян. Тогда тот «прорыв» низких частот на север, который разделяет на две части Русский Север, может являться следом низовской колонизации - в ходе борьбы Новгорода и низовских земель за северные территории. Напомним, что такой след М. В. Витов прослеживал по данным антропологии (см. раздел 4.4). Однако, хотя по группам крови АВ0 изучено то же число популяций (К = 182), что и по данным антропологии (К = 180), но генетические данные все равно остаются менее информативными из-за их неравномерного распределения по ареалу - «то пусто, то густо». Достаточно сравнить карту АВ0*0 (рис. 5.2.19.) с любой антропологической картой (например. рис. 4.2.1), чтобы убедиться, насколько антропологами лучше спланировано изучение того же числа русских популяций: они равномерно - в соответствии с историей расселения и плотностью населения - покрывают ареал, в то время как по данным генетики многие важные части ареала не обеспечены информацией. Из этого можно сделать два вывода. Во-первых, нельзя по имеющимся сейчас для русских популяций данным генетики определить точный маршрут генов - его можно набросать лишь очень приблизительно. А во-вторых, для изучения истории русского генофонда необходимо продолжать накопление данных даже по столь изученным классическим маркёрам, как группы крови АВ0! «РУССКИИ» ЛАНДШАФТ АВ0*В (рис. 5.2.22) Карта распространенности аллеля АВ0*В дополняет две предыдущие карты (частоты аллелей, показанные на всех трёх картах, в любой конкретной точке в сумме составляют единицу). Однако четкие тренды в генетическом ландшафте АВ0*В отсутствуют - они как бы исчерпаны предыдущими картами. Основная территория занята средними значениями, а на периферии разбросаны ядра высоких значений (до q = 0.43). Низкие значения видны в Псковских землях и на северо-востоке Архангельской области (где аллель АВ0*В практически отсутствует, опускаясь до q = 0.01). Однако эти два провала частот АВ0*В разделены северным хребтом высоких значений. Иными словами, мы вновь, как и для RH*d (рис. 5.2.16), видим ландшафт «лоскутного одеяла». Напомним, что на евразийских картах именно этот аллель показал четкий тренд изменения частоты с запада на восток. Похожий тренд выявлен и по аллелю RH*d. Генофонд Восточной Европы по обоим маркёрам (рис. 5.2.15. и 5.2.18) играл роль переходной зоны и обнаруживал четкий тренд «запад-восток». Однако, перейдя с регионального уровня популяционной иерархии (Восточная Европа) на этнический уровень («русские» карты), мы не обнаруживаем в русском генофонде ни такого тренда, ни соответствующей ему связи с долготой. И по RH*d (r = 0.05), и по АВ0*В (r = 0.08) корреляция с долготой близка к нулевой отметке. А сами карты набросаны вольными мазками разных «красок» значений признака. ИТОГ Обе карты - и RH*d, и АВ0*В - обеспечены обширной исходной информацией, и потому позволяют сделать некоторое обобщение: четкая закономерность на региональном уровне не «обязывает» этносы к её «исполнению». В ладу с собственной историей этнический генофонд может её «отменять» или преобразовывать в свою собственную версию генетического ландшафта. Все три аллеля локуса АВ0 вносят свой вклад в главный сценарий архитектоники русского генофонда. Эти черты мы обнаруживаем, анализируя корреляции карт аллелей локуса АВ0 с картами главных компонент русского генофонда (табл. 5.3.3). Карта аллеля АВ0*А обнаруживают высокую корреляцию (r = 0.64) с первым сценарием изменчивости классических маркёров. Вклад двух других аллелей меньше: для аллеля АВ0*0 корреляция r = 0.42, для аллеля АВ0*В корреляция r = 0.31. 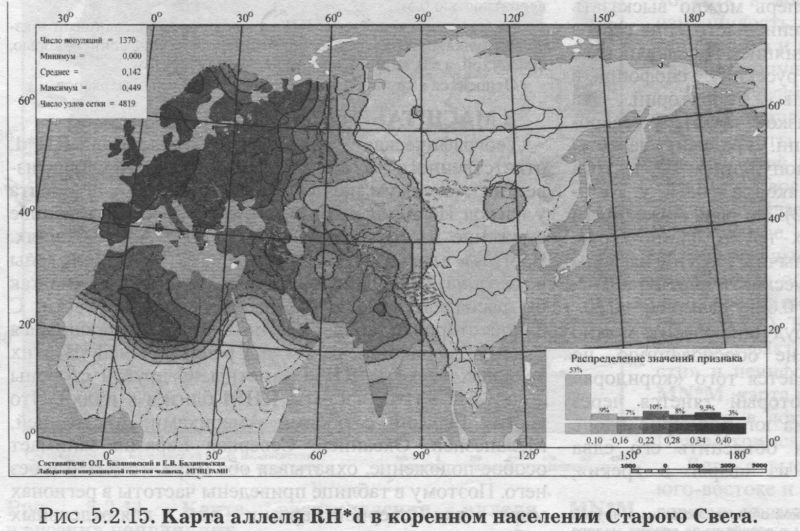
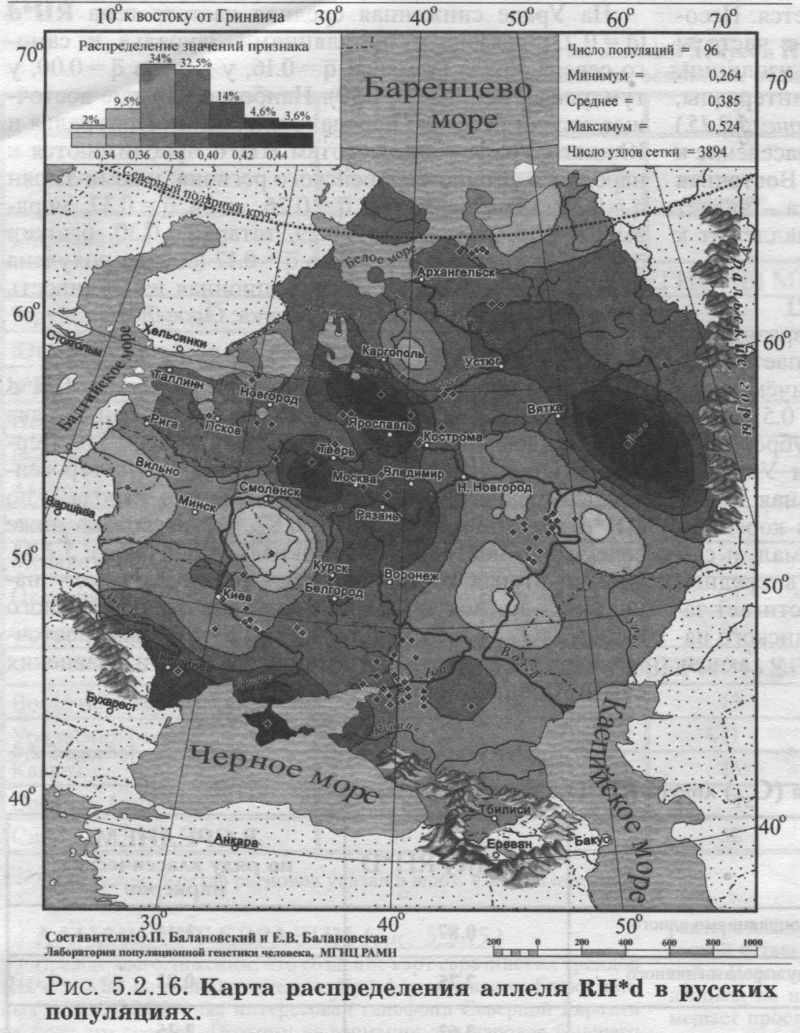
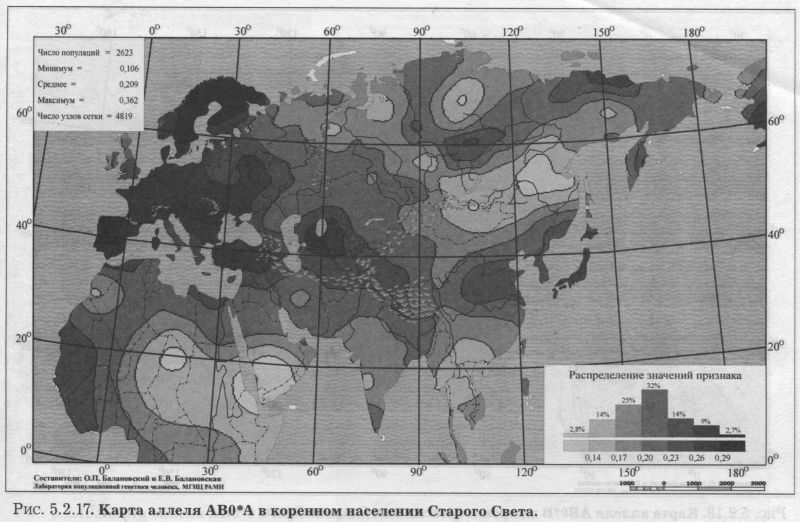
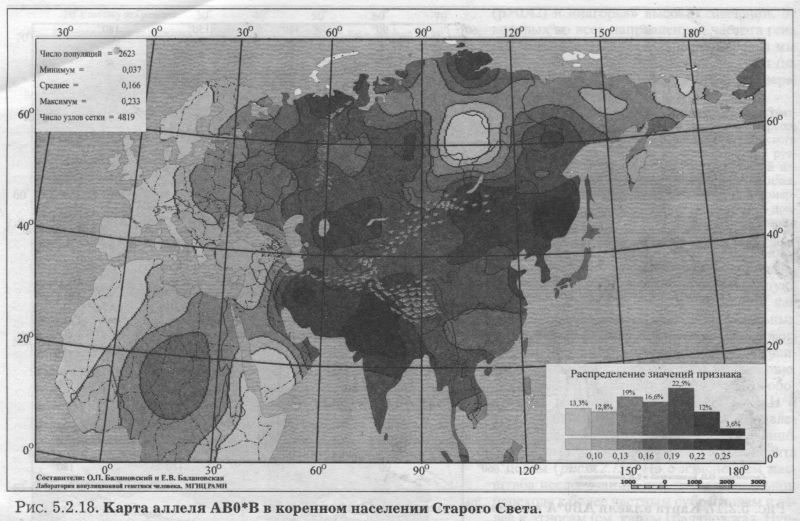
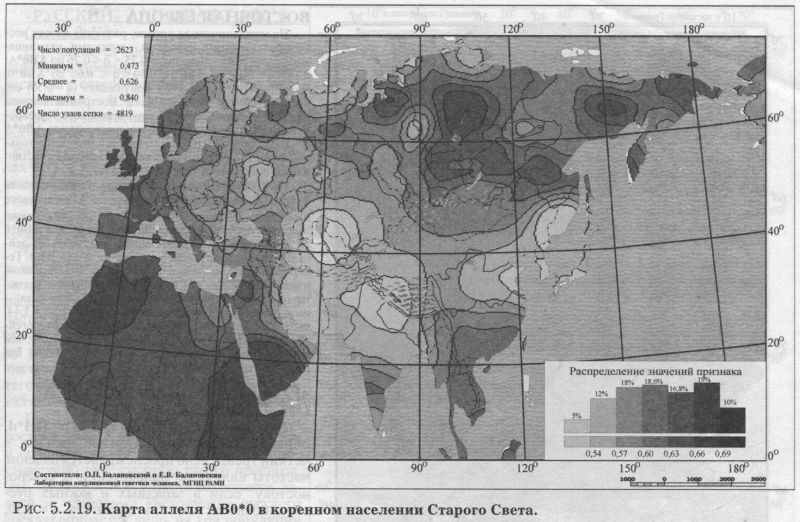
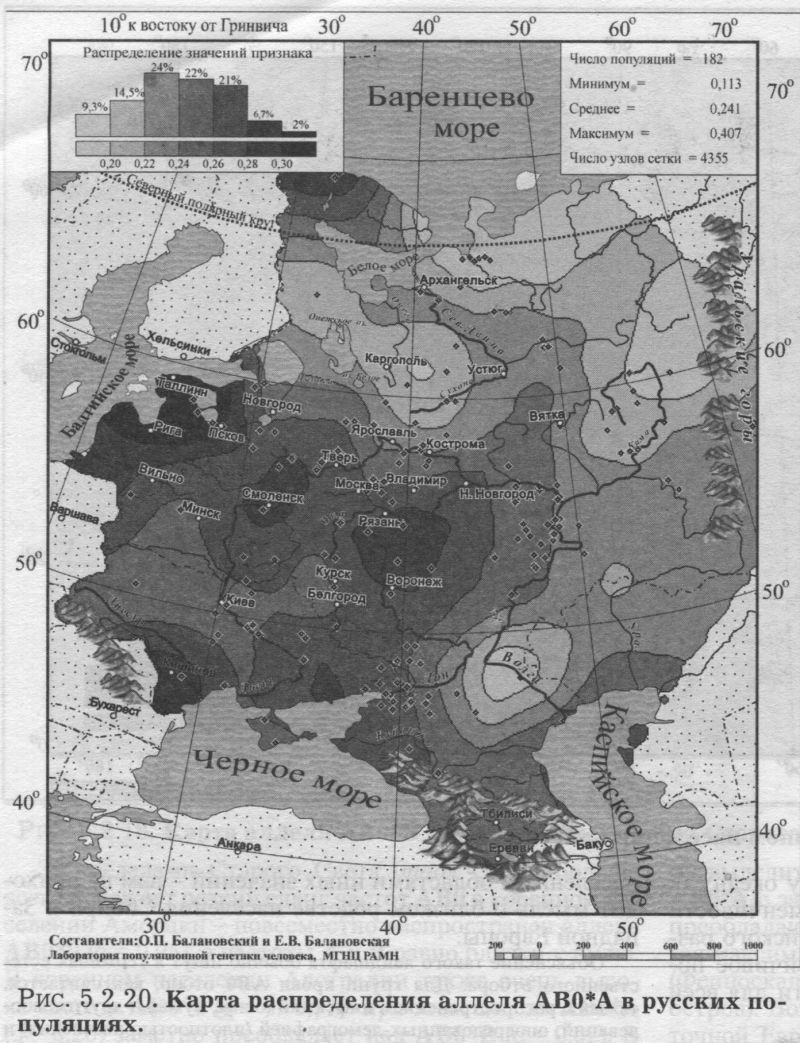
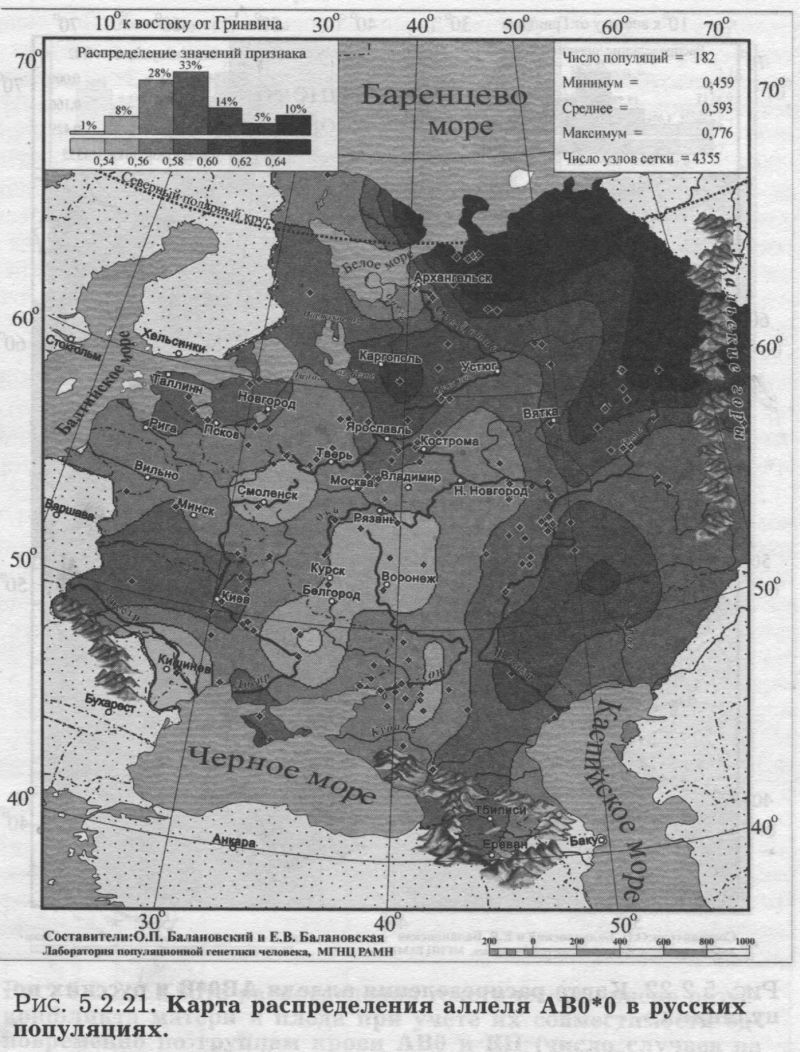
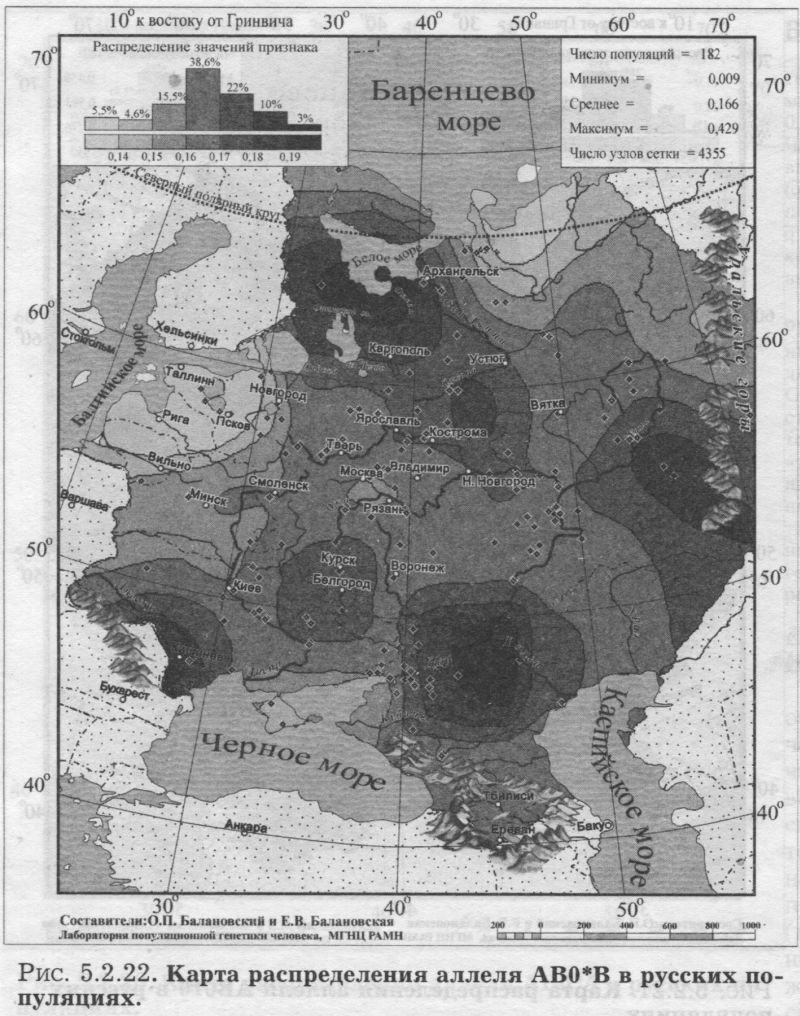
§3. Риск несовместимости матери и плода по группам крови Раз уж мы позволили себе столь подробно остановиться на изменчивости двух самых популярных классических маркёров - групп крови АВ0 и Резус, то нельзя не затронуть и их взаимодействие. Мы рассмотрим то взаимодействие, которое обычно остается за рамками самых важных «практических» применений групп крови. Мы рассмотрим генетический риск возникновения гемолитической болезни новорожденных при учете совместимости матери и ребенка не только «по резус-фактору», а одновременно по обеим системам групп крови (АВО и RH). При совместном рассмотрении обнаруживаются неожиданные и важные явления. Патология беременности, возникающая при несовместимости матери и плода по группам крови резус, общеизвестна. Проявления её могут быть очень тяжелы, и выражаться не только в анемии новорожденного, но и во множественных очагах кровоизлияний, водянке полостей тела, поражениях дыхательных путей, желудочно-кишечного тракта, печени и селезёнки [Прокоп, Гелер, 1991]. Менее известна патология при несовместимости матери и плода по группам крови АВ0. Проявления её различны на разных стадиях развития плода. В первом триместре беременности она влечет за собой увеличение частоты внутриутробной гибели эмбриона или плода. Потери из-за элиминации АВО-несовместимого с матерью потомства весьма ощутимы: они составляют по разным оценкам от 3-5 % до 15-21% [Туманов, Томилин, 1969; Головачев, 1983]. В конце последнего триместра беременности, перед родами, происходит переход антител и других факторов гуморального иммунитета через плаценту. Такая пассивная иммунизация имеет важнейшее значение для новорожденного, поскольку компенсирует незрелость иммунных систем ребенка [Головачев, 1983]. Побочными эффектами такой иммунизации может быть гемолитическая болезнь и ухудшение функционального состояния новорожденного, связанные с иммунной несовместимостью [Туманов, Томилин, 1969; Головачев, 1983]. Последствия несовместимости по системе АВО наблюдаются в 2-3 раза чаще, чем по другим иммунным системам [Доссе, 1959]. Но они, в отличие от резус-несовместимости, проявляют себя раньше - до появления ребенка на свет. Для плода они могут иметь печальные последствия. А для новорожденного серьезной угрозы они обычно не представляют, следовательно, не представляют они и проблемы для здравоохранения. Логично было бы ожидать, что в случаях двойной несовместимости матери и плода - по АВ0 и по Резус - угроза для плода резко возрастёт. Однако на самом деле все наоборот. И число случаев, и интенсивность патологии при двойной несовместимости - по АВО и по Резус - резко снижаются [Доссе, 1959; Соловьева, 1964; Туманов, Томилин, 1969; Головачев, 1983]. Причина этому - «конкуренция антител». Эритроциты плода, попадая в организм матери, разрушаются под влиянием уже присутствующих в её крови АВ0-антител, не успевая вызвать иммунизацию к RH-антигену. Поэтому в популяциях наибольший риск возникновения иммунного резус-конфликта, значимого с точки зрения здравоохранения, возникает при несовместимости матери и ребенка по RH, сопровождающейся совместимостью по АВ0. То есть при резус-отрицательности матери (RH-) и резус-положительности (RH+) плода наибольшая угроза гемолитической болезни новорожденного возникает тогда, когда группы крови АВ0 одинаковы у матери и ребенка: например, А и А, В и В. 0 и 0. Такое сочетание групп крови АВ0 и Резус составляет, по разным оценкам, от 66 % [Соловьева. 1964] до 90 % [Michoud, 1961] всех случаев гемолитической болезни, вызванной резус-несовместимостью матери и плода. Построив карту, отражающую вероятность того, что у резус-отрицательных матерей с группами крови А, В и 0, будут резус-положительные дети, обладающие группами крови АВ0, идентичными материнским, мы картографируем распространение основной группы генетически обусловленного риска возникновения резус-несовместимости матери и плода. Мы не раз говорили, что карта является не только графическим, но и алгебраическим объектом, не только «картинкой», но и числовой матрицей, с которой можно производить любые необходимые преобразования. Мы постоянно пользуемся этим неоспоримым достоинством наших карт. Их «сглаживание», корреляция, расчёт главных компонент и генетических расстояний - все эти процедуры включают порой очень длинную цепь преобразований, которые мы совершаем с исходными картами. Покажем эту цепь на примере карты генетического риска: для создания единственной результирующей карты (рис. 5.2.23.) мы построили 115 промежуточных карт. В табл. 5.2.6. указаны типы браков, в которых возможно появление детей, идентичных матери по группе крови А, В или 0, но с угрозой возникновения RH- конфликта. Чтобы получить карту генетического риска несовместимости матери и плода, были проведены следующие преобразования карт: 1) Получены карты распространения частот аллелей локусов АВ0 и RH (пять карт пяти аллелей); 2) Получены карты распространения генотипов родителей (17 карт, генотипы указаны в табл. 5.2.6.) 3) Перемножая карты родительских генотипов, получаем карты частот указанных типов браков. Сочетания генотипов с нулевой вероятностью возникновения резус-конфликта мы не рассматриваем и карты для них не строим. Для оставшихся 46 сочетаний родительских генотипов мы строим 46 промежуточных карт. 4) Умножив эти карты на вероятность, указанную в табл. 5.2.6., получаем вероятность возникновения RH-несовместимости в каждом из этих браков (ещё 46 карт). 5) Сложением этих карт получаем суммарную карту, которую умножаем на 1/3 (считается, что генетически эффективная часть популяции составляет около 1/3 от её численности). 6) Умножив результирующую карту на 100 ООО, получим карту генетического риска возникновения RH-конфликта матери и плода, выраженную в числе случаев на 100 ООО населения. Полученная таким образом карта генетического риска резус-конфликта матери и плода при учете групп крови АВ0 и Резус дана на рис. 5.2.23. Карту для коренного населения всей Северной Евразии мы не приводим, так как в азиатской части частота RH*d близка к нулю и генетический риск минимален. Лишь некоторое его увеличение (около 500 случаев на 100 000) прогнозируется вдоль северной границы Казахстана и в среднеазиатских республиках. Однако в европейских районах с повышенной концентрацией гена RH*d величина генетического риска достигает весьма значимых величин (рис. 5.2.23.). В отдельных районах России, Белоруссии, Литвы и Грузии величина генетического риска резус-несовместимости матери и плода, угрожающей здоровью ребенка, составляет около 2000 случаев на 100 000 населения. Однако все не так страшно. Обратим внимание, что мы говорим только о генетическом риске резус-конфликта. Дело в том, что генетический риск и риск развития гемолитической болезни новорожденного - это две разные вещи. Даже при наличии фактора несовместимости (генетический риск), гемолитическая болезнь возникает лишь у части детей из группы риска - примерно у 15 % (наш специальный расчёт по совокупности литературных источников). Будь доля больных детей постоянной во всех популяциях, география заболеваемости полностью повторяла бы географию генетического риска. Однако вариабельность этой доли велика. Поэтому география заболеваемости гемолитической болезнью новорожденных начинает зависеть не только от генетического риска, но и средовой компоненты - как внешней (демографической, экологической, социальной), так и внутренней (например, индивидуальный уровень гетерозиготности организма или даже резус-фактор бабушки новорожденного, повлиявший на иммунизацию матери в период её внутриутробного развития). Сопоставление географии наблюдаемой заболеваемости с географией генетического риска и географией гетерозиготности позволяет прогнозировать результативность профилактических мероприятий и соответственно их корректировать. 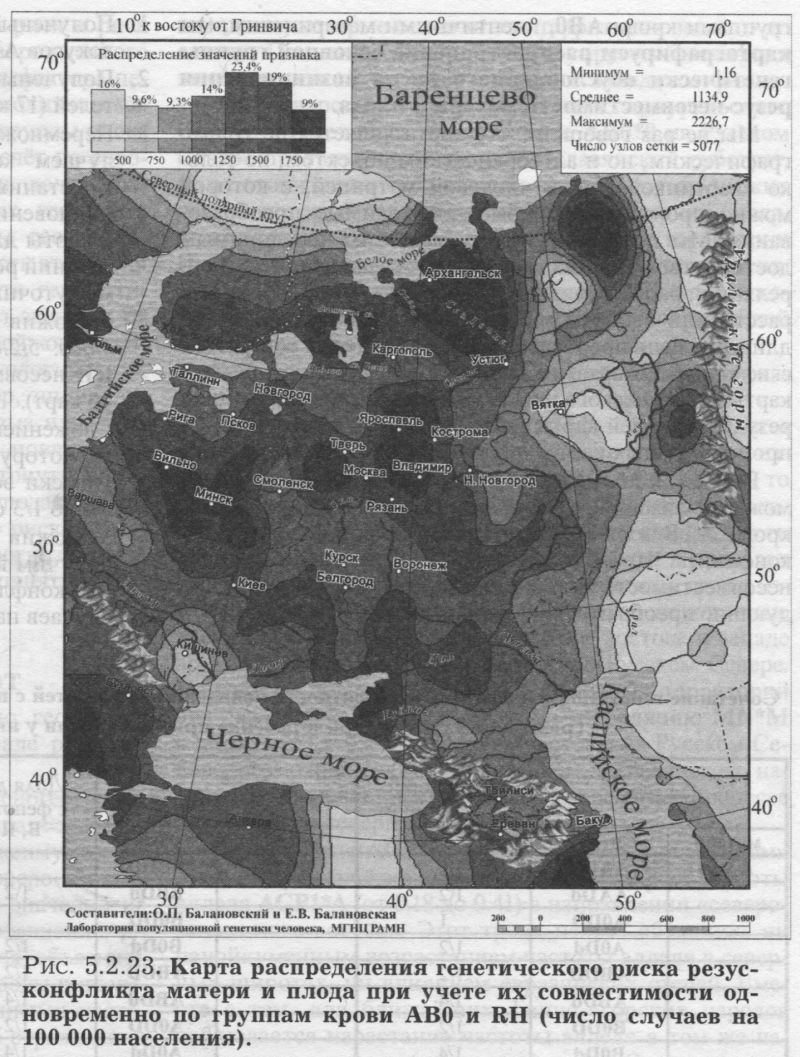
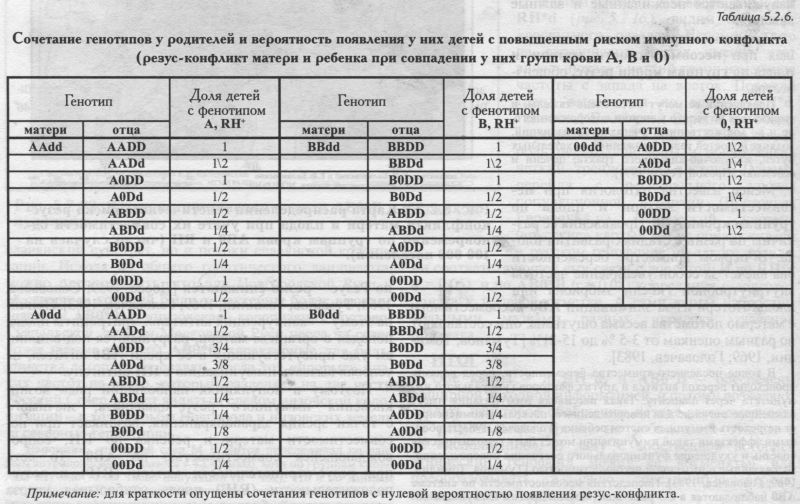
§4. Обзор «простых» карт Подведем краткий итог рассмотрению географии отдельных классических маркёров в ареале русского народа. Яркая закономерность выявляется геногеографической картой НР*1: чёткий тренд «юго-запад<=>северо-восток». Клинальная изменчивость с максимумом на юге q = 0.51 и снижением частоты к северо-востоку q = 0.17 нарушается зоной среднерусских значений на арктическом побережье (q = 0.36). Это повышение частоты НР*1 в русских популяциях нельзя объяснить влиянием коренных народов Арктики - народы циркумполярной зоны характеризуются пониженными частотами НР*1. Можно выдвинуть гипотезу об отражении в генетическом ландшафте НР*1 этапа освоения северных окраин Руси новгородцами в XI-XIV веках. Не менее чёткий тренд наблюдается и в географической изменчивости АВ0*А в русских популяциях: мы вновь видим падение частоты аллеля по направлению к северо-востоку. Несмотря на небольшой размах изменчивости АВ0*А, практически всё множество популяций соответствует картине плавного снижения частоты к востоку и северу, тем самым подтверждая объективность этой закономерности. При этом генетический ландшафт АВ0*А не только русского народа, но и генофонда Восточной Европы своеобразен и необъясним в терминах «переходной зоны» между Европой и Азией. Генетический ландшафт русского генофонда по АВ0*0 демонстрирует тот же тренд, но является почти зеркальным отражением АВ0*А - с северо-востока на юг и юго-запад идет не увеличение, а падение частоты. И вновь, несмотря на небольшой размах изменчивости, почти все популяции следуют рельефу плавного снижения частоты с севера к юго-западу. Сравнение с окружающими народами не исключает, что повышение частоты АВ0*0 В северных русских популяциях связано с финно-угорским субстратом - теми племенами и народами, которые освоили эти территории задолго до славян. По данному гену реконструируемый генофонд этих популяций тяготеет больше к западным финно-язычным народам, чем к восточным, приуральским. Однако в генетическом ландшафте АВ0*0 на Русском Севере проявляется не только дославянский субстрат, но и потоки славянской колонизации. «Прорыв» низких частот, который разделяет на две части Русский Север, может являться следом борьбы за северные территории Новгорода и низовских земель - ростово-суздальских, а затем московских. Возможно, именно их продвижение на север отражено на карте АВ0*0, сохранившей следы этих генных потоков. По локусу MN различия между всеми популяциями Восточной Европы (GST = 1.2) рамного меньше селективно-нейтрального уровня (GST = 2.6). При этом гетерогенность русского народа (GST = 1.2) равна изменчивости всех народов Восточной Европы по MN, то есть охватывает изменчивость всего коренного населения региона. При полной «безликости» и одинаковости этнических частот MN*M по всей Европе, в пределах русского генофонда обнаруживается четкий тренд изменчивости. Рельеф задают полярно расположенные элементы - ядра максимумов на юго-востоке и западе ареала, область низких значений на Русском Севере. При почти полной аморфности основных территорий эти два полюса создают высокую корреляцию MN*M с широтой: r = 0.44. Снижение частоты на Русском Севере, возможно, связано с влиянием дославянского населения, включённого в русский генофонд в процессе колонизации северных территорий. Карта аллеля АСР1*А демонстрирует в «исконном» русском ареале чётко выраженный тренд: рост частоты аллеля АСР1*А (от 0.18 до 0.41) в направлении «северо- запад<=>юго-восток». Этот тренд нельзя объяснить ни панойкуменным возрастанием частоты аллеля в северных широтах, ни влиянием евразийских степей. Вместе с тем, для ближайших финно-угорских народов наблюдается нарастание частоты аллеля в том же направлении: с севера (и с запада) на юг. Это позволило выдвинуть гипотезу об отражении в «русском» тренде АСР1*А финно-угорского субстрата - дославянского населения Русской равнины. Карта другого аллеля этого локуса - АСР1*С - при средней частоте аллеля в русском ареале q = 0.03 выявляет общее снижение частоты к северу от Волго-Окского междуречья (до 1-2%) и повышение частоты АСР1*С в центральных, восточных и южных областях (до 7%). Однако при столь низких частотах требуются много большие выборки и большее число изученных популяций. Это замечание относится и к карте низко полиморфного аллеля PGD*C (0.02 < q < 0.06, q = 0.04), который к тому же, по-видимому, находится под давлением стабилизирующего отбора (GST = 0.35, то есть в 4 раза ниже селективно-нейтрального уровня GST = 1.36). Для аллеля РI*М2 характерна иная - не широтная, а долготная изменчивость, реже наблюдаемая в русском генофонде. На карте распространения аллеля РI*М2 в «исконном» русском ареале видна хорошо выраженная клинальная изменчивость: падение частоты в направлении «запад<=>восток» от q = 0.23 до q = 0.04. Корреляция частоты РI*М2 с долготой достигает значений r = 0.9. Этот тренд совпадает с градиентом частот в Северной Евразии, где частота аллеля в целом убывает в направлении с запада на восток. Карты других генетических систем не выявляют столь чётких трендов. Однако и для них можно выделить некоторые особенности. Так, для генетического ландшафта аллеля GC*2 структурообразующим элементом является «впадина» (перепад значений от 0.38 до 0.14), следующая вдоль сорокового меридиана (40° в.д.). Для карты аллеля GC*1S характерна широтная изменчивость с повышением частоты аллеля от 0.48 на северо-западе и северо-востоке русского ареала до q = 0.68 к югу от 55 параллели (55° с.ш.). Своеобразен генетический ландшафт аллеля TF*C2. Для него характерны три широтных зоны: северная - повышенных частот (q = 0.21) , центральная - пониженных частот (q = 0.06), южная - со среднерусскими частотами (q = 0.15). Карты всех аллелей локуса PGM1 являются мозаичными и не поддаются интерпретации, так что информативность локуса PGM1 для изучения русского генофонда невелика. При этом средний по локусу PGM1 уровень межпопуляционной изменчивости (GST = 0.75) в два раза меньше селективно-нейтрального уровня (GST = 1.36), что указывает на возможное действие стабилизирующего отбора. Лишь в генетическом рельефе аллеля PGM1*1- всё же можно предположить некоторую закономерность. В целом средние (q = 0.10) и низкие частоты (q = 0.00) PGM1*1- характерны для западных (новгородско-псковских) и волго-окских популяций. Высокие (q = 0.18) частоты PGM1*1- наблюдаются к северу и востоку - в вологодских и костромской популяциях, вновь снижаясь на крайнем севере. В генетическом рельефе ESD*1 мы снова видим чередование четырёх широтно расположенных волн (с колебанием частот от 0.79 до 0.94). При этом впадину низких значений на юго-востоке русского ареала нельзя объяснить влиянием соседних тюркоязычных народов, поскольку у них частота аллеля достаточно велика. Карта аллеля GL01*1 ещё своеобразнее: центральная зона занята высокими значениями частоты аллеля (q = 0.48), а к северу и к югу частота аллеля снижается до минимальных значений (q = 0.27). Причём такой ландшафт прямо противоположен ожидаемому из характеристик соседних народов. ВЫВОДЫ Из серии карт изменчивости отдельных генов в ареале русского генофонда удается сделать лишь два общих вывода. Во-первых, карты классических генных маркёров являют довольно сложные ландшафты, среди которых можно всё же - с определённой степенью достоверности - выделить широтную изменчивость. Хотя при этом непрерывная клинальная изменчивость прослеживается на картах лишь нескольких аллелей - НР*1, АСР1*А, MN*M, АВ0*А, АВ0*0. Широтное направление в целом соответствует географии диалектов и определённым историческим этапам - колонизации северных окраин Руси, освоения степи и др. Вторым, но более интересным результатом анализа отдельных генов является тот факт, что повышение или понижение частоты какого-либо аллеля на границах русского ареала в большинстве случаев нельзя объяснить обменом генами с соседними народами. Например, частоты аллеля MN*M в русских популяциях, формирующие ядра высоких значений, нельзя однозначно вывести из частот в популяциях окружающих их народов. И снижение частоты АСР*А у русских Поволжья не объясняется смешением со степными народами, поскольку у последних частота этого аллеля повышена. Мы приводили и другие примеры. Это служит указанием на большую, чем можно было предположить, роль не смешений, а внутриэтнических процессов в формировании русского генофонда (возможно, основным фактором может быть дрейф генов в окраинных популяциях). Картографирование частот аллелей позволило выявить клинальную изменчивость в русском генофонде для ряда аллелей. Однако для подавляющего большинства изученных генов клинальная изменчивость не обнаружена. В целом, мы видим ряд чрезвычайно разнообразных генетических ландшафтов - долготный тренд, «диагональная» изменчивость, широтно расположенные волны колебания частоты аллеля, «лоскутное одеяло». Какова роль того или иного ландшафта в русском генофонде, покажет дальнейший анализ обобщённых карт (главных компонент и генетических расстояний). Однако уже на этом этапе можно говорить о своеобразии русского генофонда, пространственную структуру которого нельзя понять лишь из знания генофонда окружающих его народов. Этот результат указывает на значительную гетерогенность и сложность формирования русского генофонда. Мы рассмотрели русский генофонд на фоне не только его соседей, но и в «глобальном» масштабе - в пространстве Северной Евразии и Евразии в целом. При этом оказалось, что, переходя от глобального рассмотрения к регионам (вложенным «матрёшкам»), мы можем обнаруживать исчезновение глобальных закономерностей. Именно это мы и обнаруживаем в генетическом ландшафте АВ0*В и RH*d. На евразийских картах RH*d и АВ0*В генофонд Восточной Европы играл роль переходной зоны и обнаруживал чёткий тренд «запад-восток». Однако перейдя с глобального (Евразия) на этнический уровень (русский ареал), мы не находим в русском генофонде такого тренда: и по RH*d, и по АВ0*В корреляции с долготой нет! На карте русского народа мы не видим даже намёка на общий трансевразийский тренд «запад-восток»! Более того, мы не видим вообще никаких чётких закономерностей. Карты подобны «лоскутному одеялу», где области высоких значений мирно соседствуют с областями низких значений. Как мы говорили, чёткая закономерность на глобальном уровне не «обязывает» этносы к её «исполнению». В ладу с собственной историей этнический генофонд может её «отменять» или преобразовывать в свою собственную версию этнического ландшафта. Итак, реальность опровергает простую схему, предполагающую, что континентальный тренд выложен, как из «кирпичиков», из этнических трендов, и каждый из этносов отражает ту часть всеобщего тренда, которая ему положена в соответствии с размером и местом его ареала. С изменением масштаба исследования, при последовательном переходе к более частным регионам и далее к этносам континентальный тренд может быть многократно перекрыт более частными событиями истории этих генофондов. Эти примеры демонстрируют важный методический принцип. Основные - глобальные - тренды формируются под действием факторов истории и природы в течение длительного времени, и для их выявления необходим соответствующий пространственно-временной масштаб. Соответствие масштаба времени действия эволюционного фактора масштабам генофонда и его географического ареала — важнейший принцип в изучении закономерностей формирования генофондов [Рычков, 1973, 1984; Рычков, Ящук (Балановская), 1980, 1983]. 5Это Восточная Европа. Кавказ. Урал, Средняя Азия, Сибирь. Специально уточним, что, согласно такому делению, регион «Восточная Европа» не включает в себя ни данные по Северному Кавказу (которые включены в регион «Кавказ»), ни по Приуралью (включены в регион «Урал»). 6Возможное действие отбора оцениваюсь при сравнении межпопуляционной изменчивости для данного аллеля и селективно-нейтральной изменчивости (см Прн.тхсепие) Селективно-нейтральный уровень изменчивости русского генофонда рассчитан в среднем по всем картам 44 аллелей 17 иммуно-биохимических локусов (табл. 5.2.1.). наиболее подробно изученных в русском генофонде Это среднее значение межпопуляционной изменчивости, рассчшаннос но картографическим моделям, составило (GST=1.36x10-2. Здесь и далее в разделе 5.2. это значение используется как оценка селективно-нейтральной изменчивости русского генофонда 7Здесь и далее в тексте указываются абсолютные значения коэффициентов корреляции без указания знака связи (взятые по модулю), поскольку при обсуждении важна сила связи, а не ос направление. Точные значения коэффициентов корреляции с указанием знака связи приведены в таблице 5.2.1 8Популяцию русских Житомирской области Украины с низкой частотой аллеля MN*M мы не принимаем в расчёт из-за крайне малого размера выборки. 9 На картах регион Восточной Европы включает и Приуралье, и Северный Кавказ. 10Напомним, что мы отобрали для картографирования все популяции, расположенные от 43° до 70° северной широты и от 24° до 60° восточной долготы. 11Следуя в определении границ «исконного» ареала за В. В. Бунаком [Происхождение.....1965]. |
загрузка...